
|
|
|
|
|---|---|---|
|
|
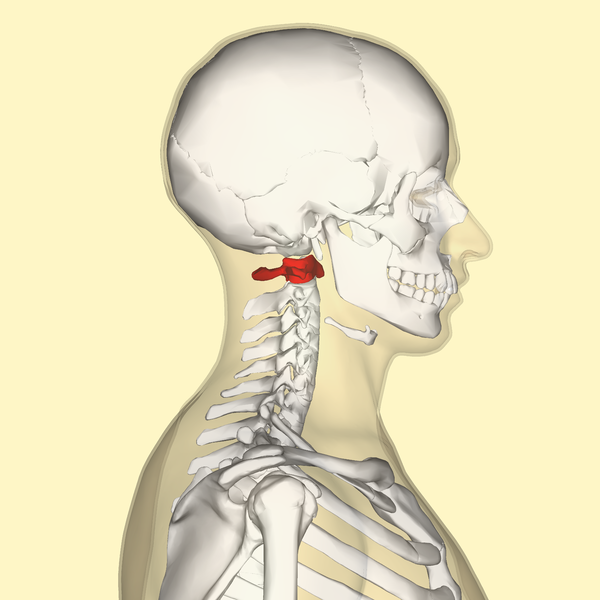
You can judge the health of a brain by observing the stability of the head and eyes. The more stable they are, the better you can think. If you watch a Shaolin monk you will see that their heads and eyes are always under control. What follows is a physics discussion of the geometry of the neck vertebrae, the brain, the ears, and the eyes, with the purpose of providing the tools required to assess mental health.
 |
 |
 |
|---|---|---|
 |
 |
 |
|---|---|---|
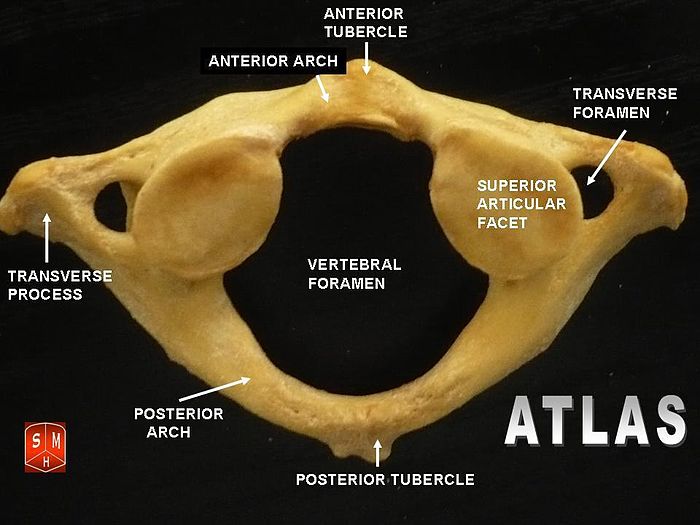
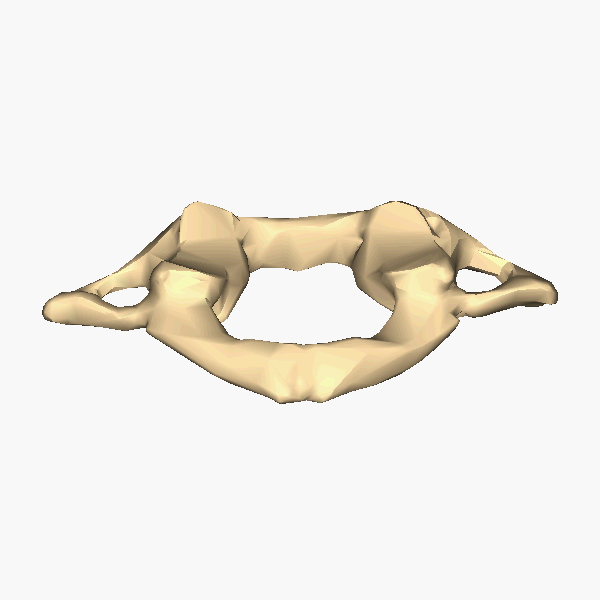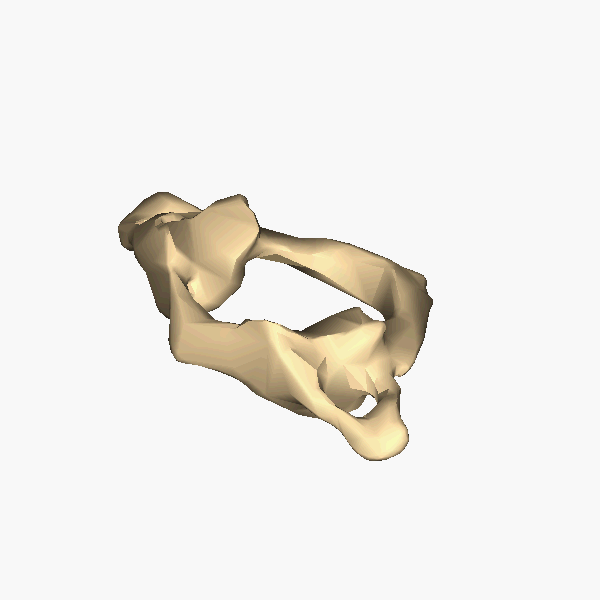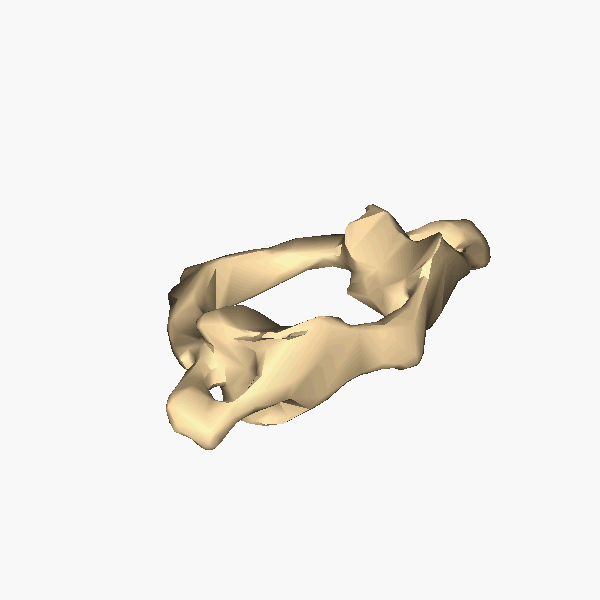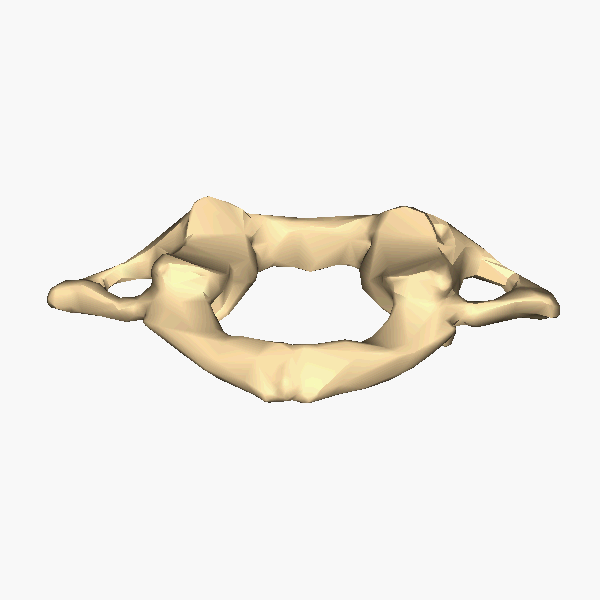
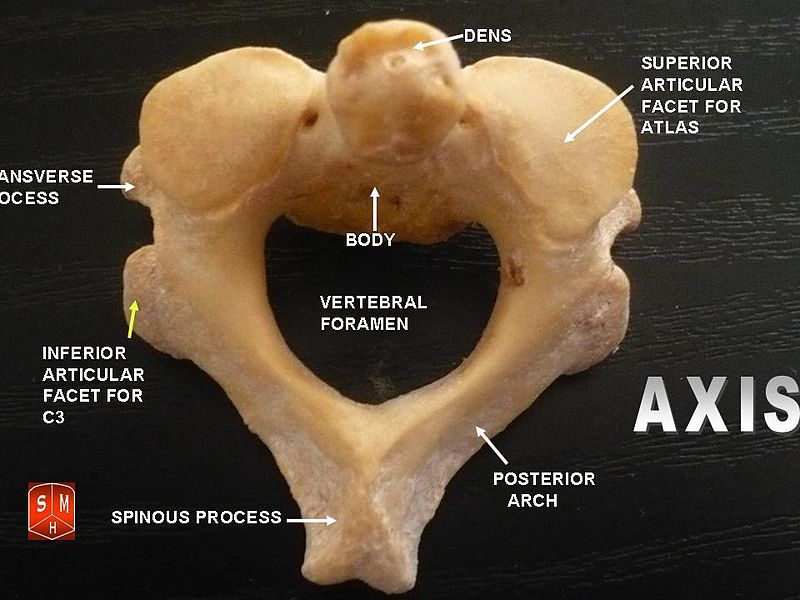
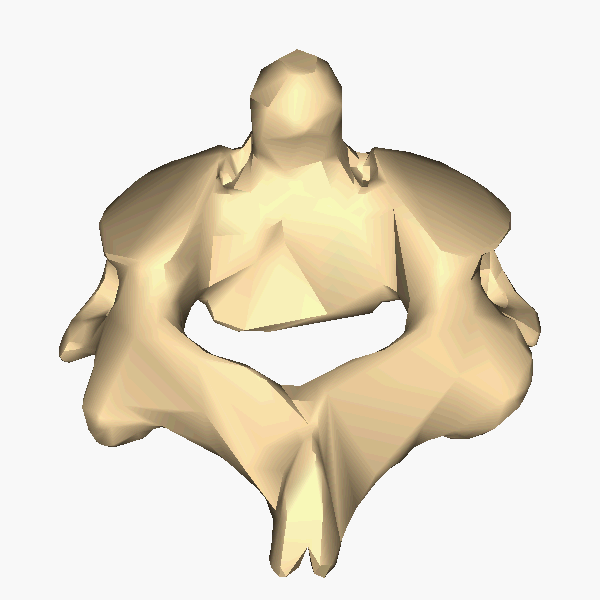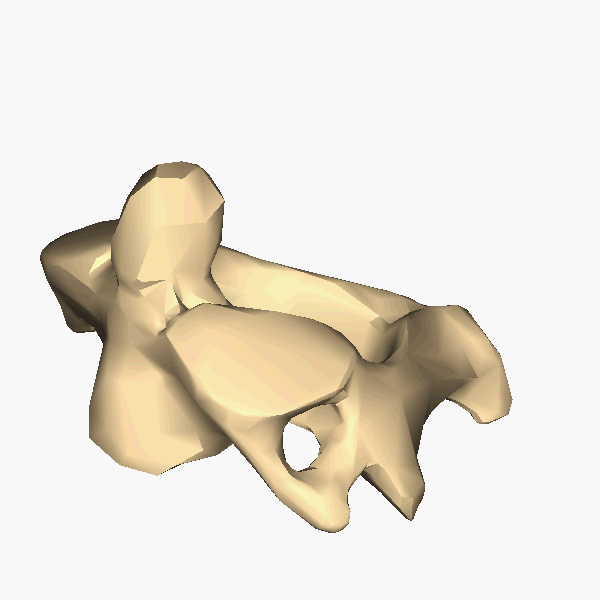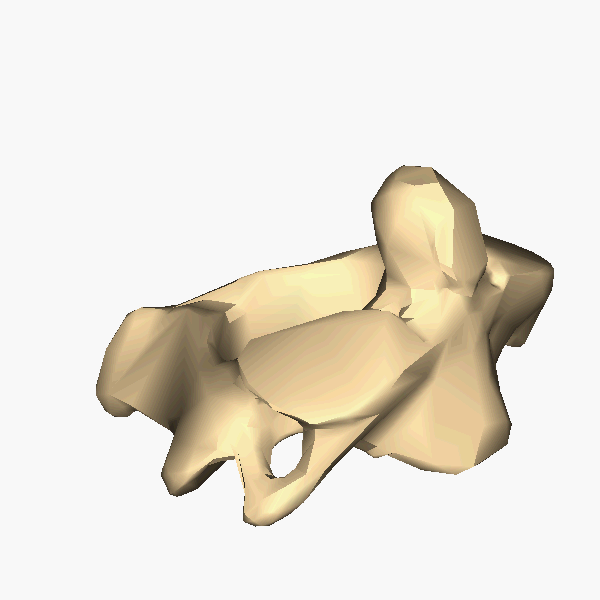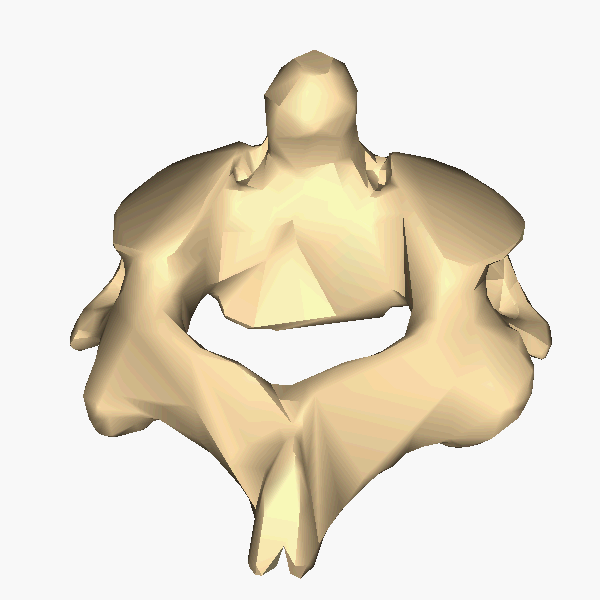
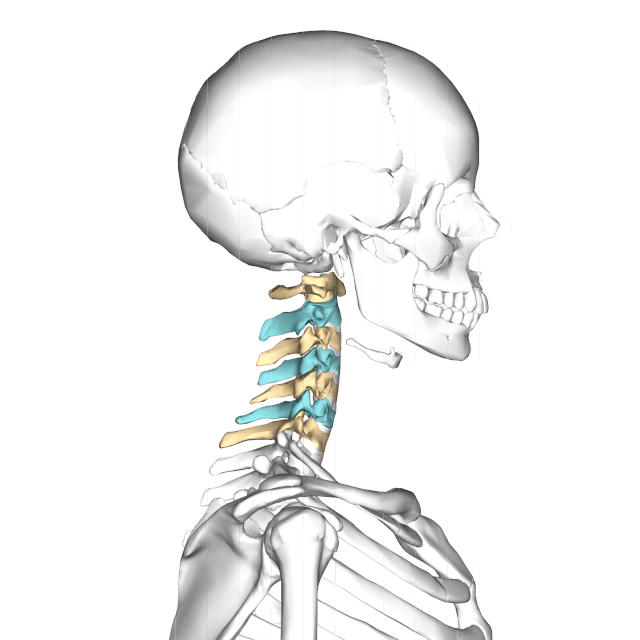
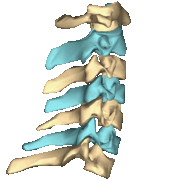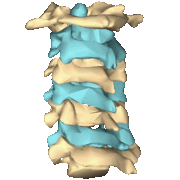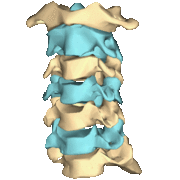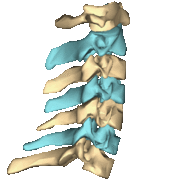
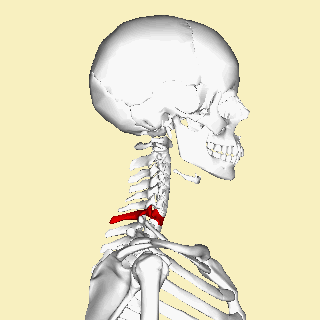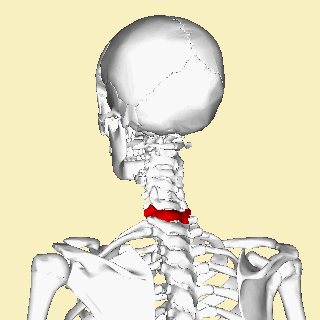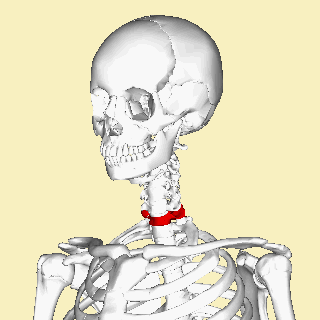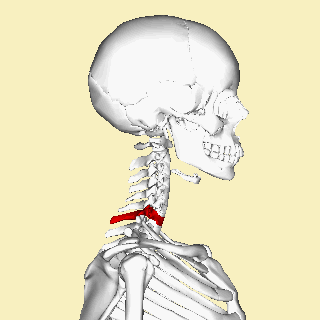
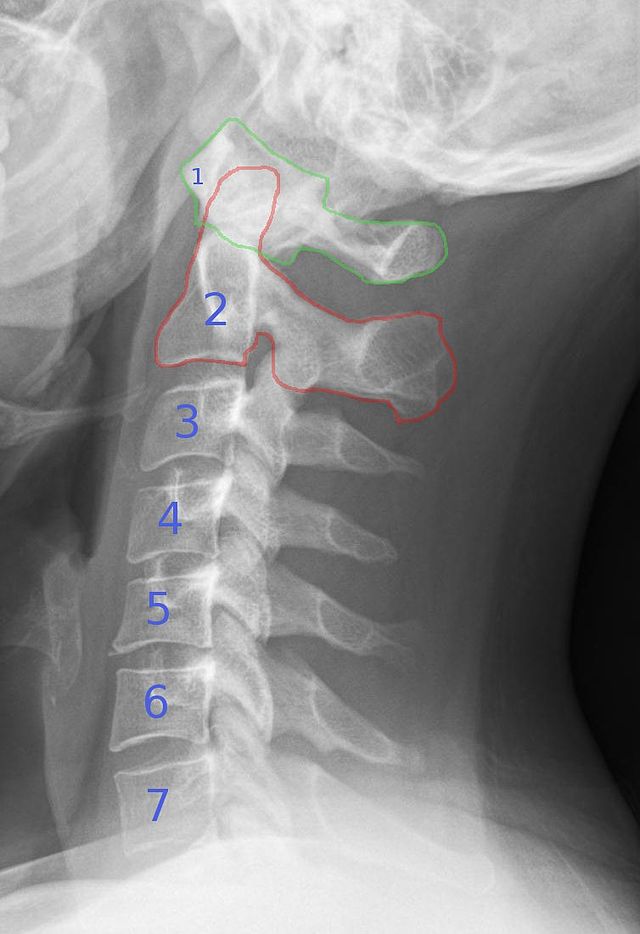
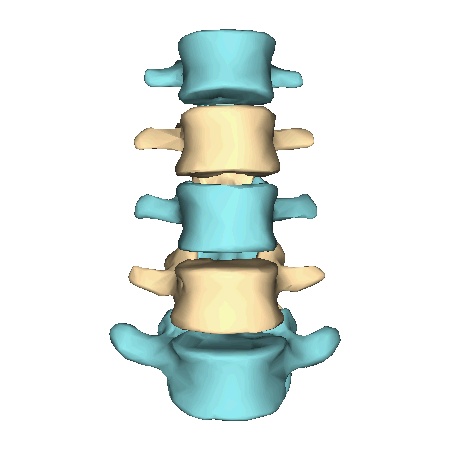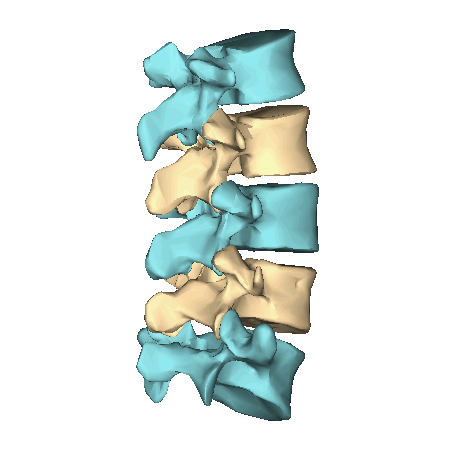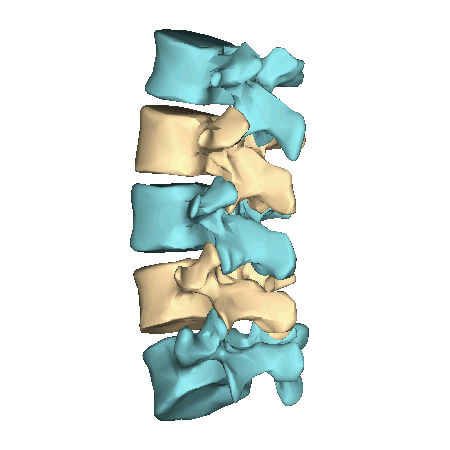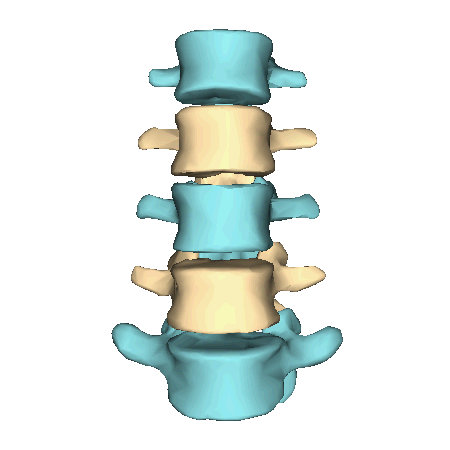
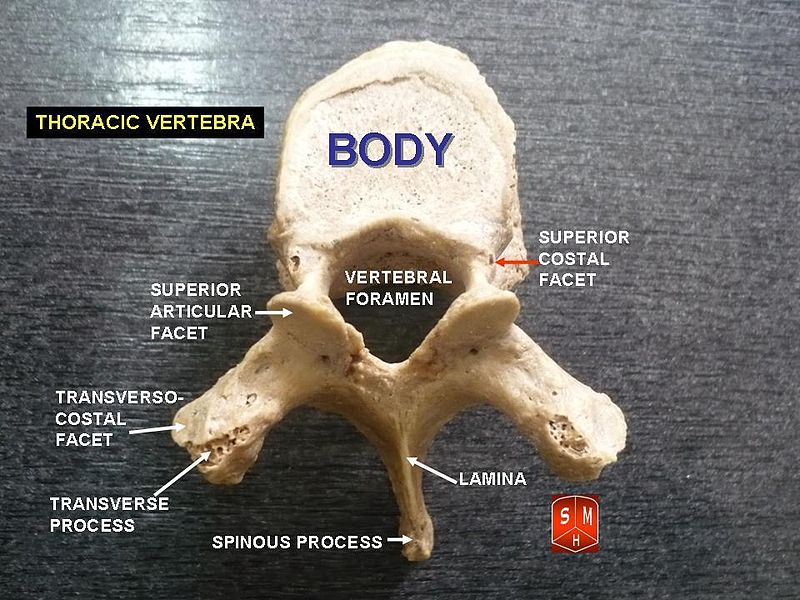
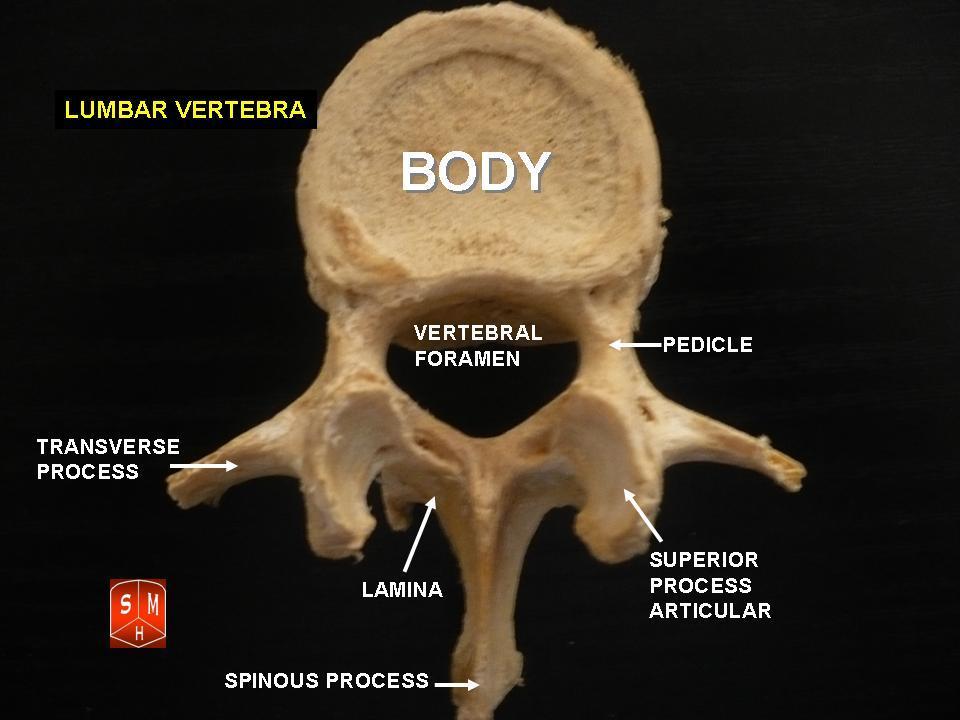
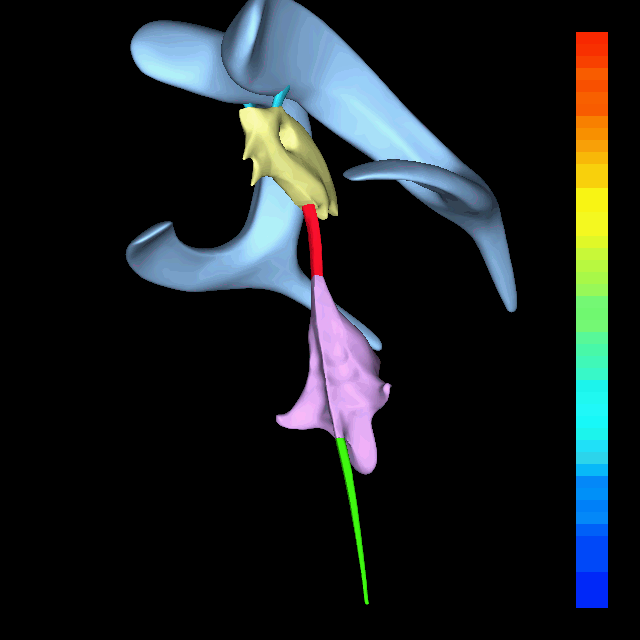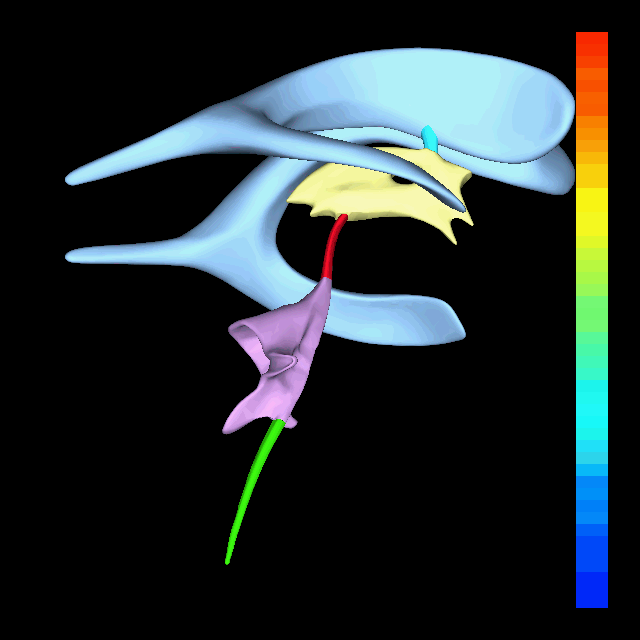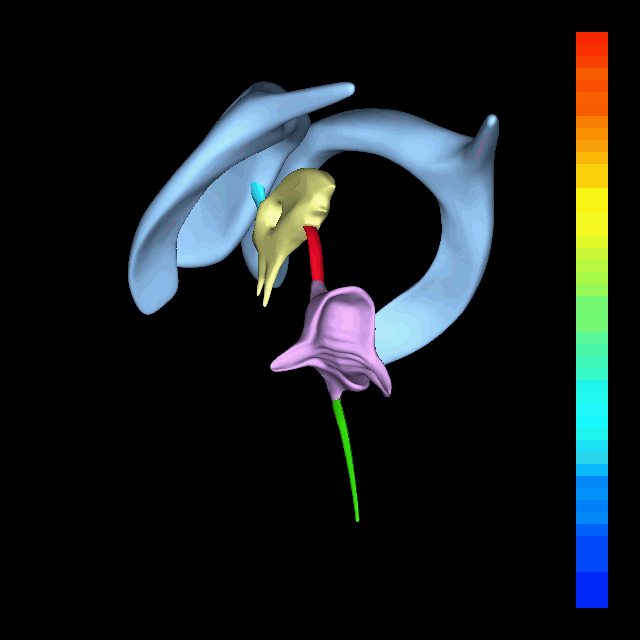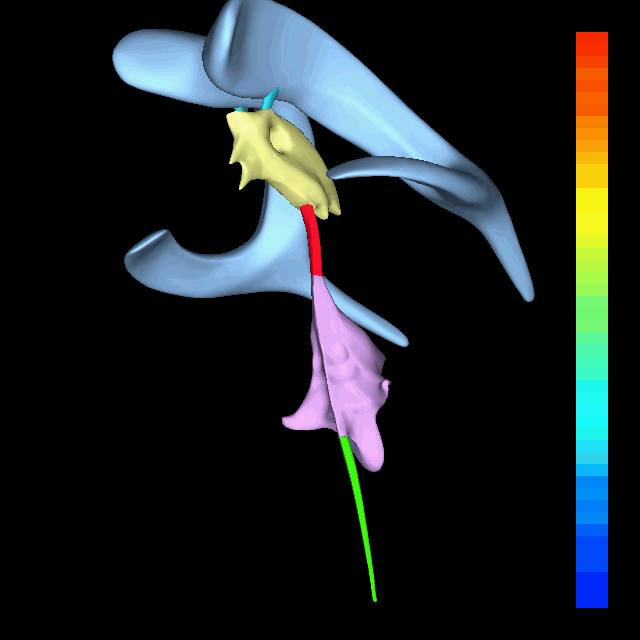
The atlas and axis vertebra move your head the same way an alt-azimuth mount moves a telescope.
 |
 |
|---|---|
 |
 |
 |
|---|---|---|
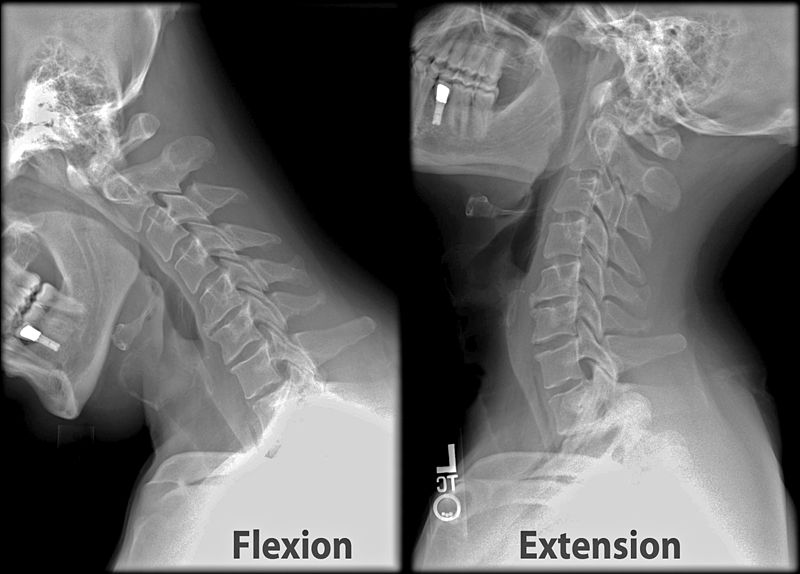
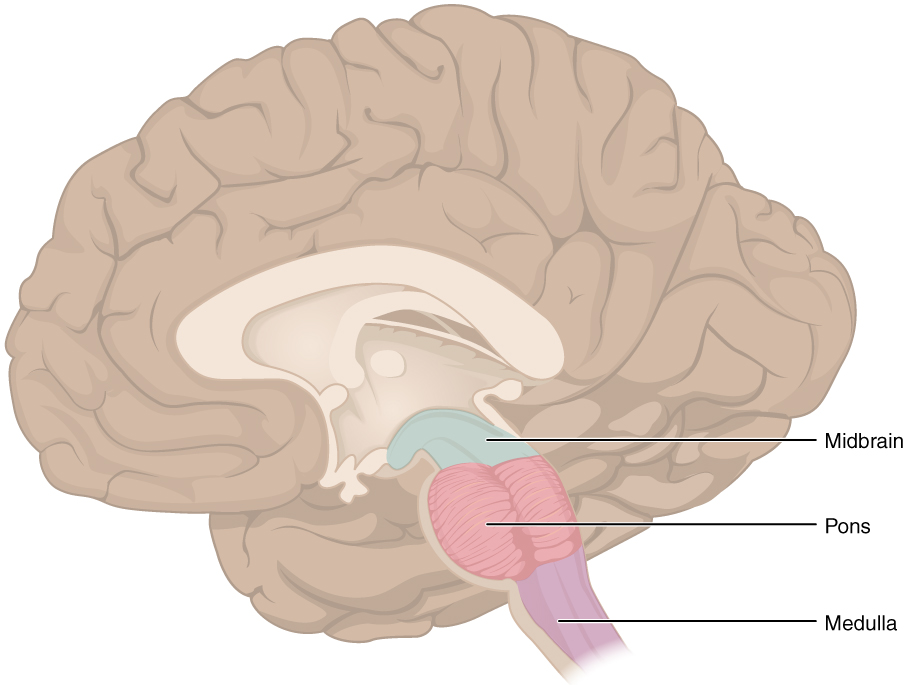
Pitch is controlled by the Atlas-Skull joint. (Nodding your head "yes")
Yaw is controlled by the Axis-Atlas joint. (Shaking your head "no")
Roll is controlled collectively by all neck vertebrae.
 |
 |
|---|---|
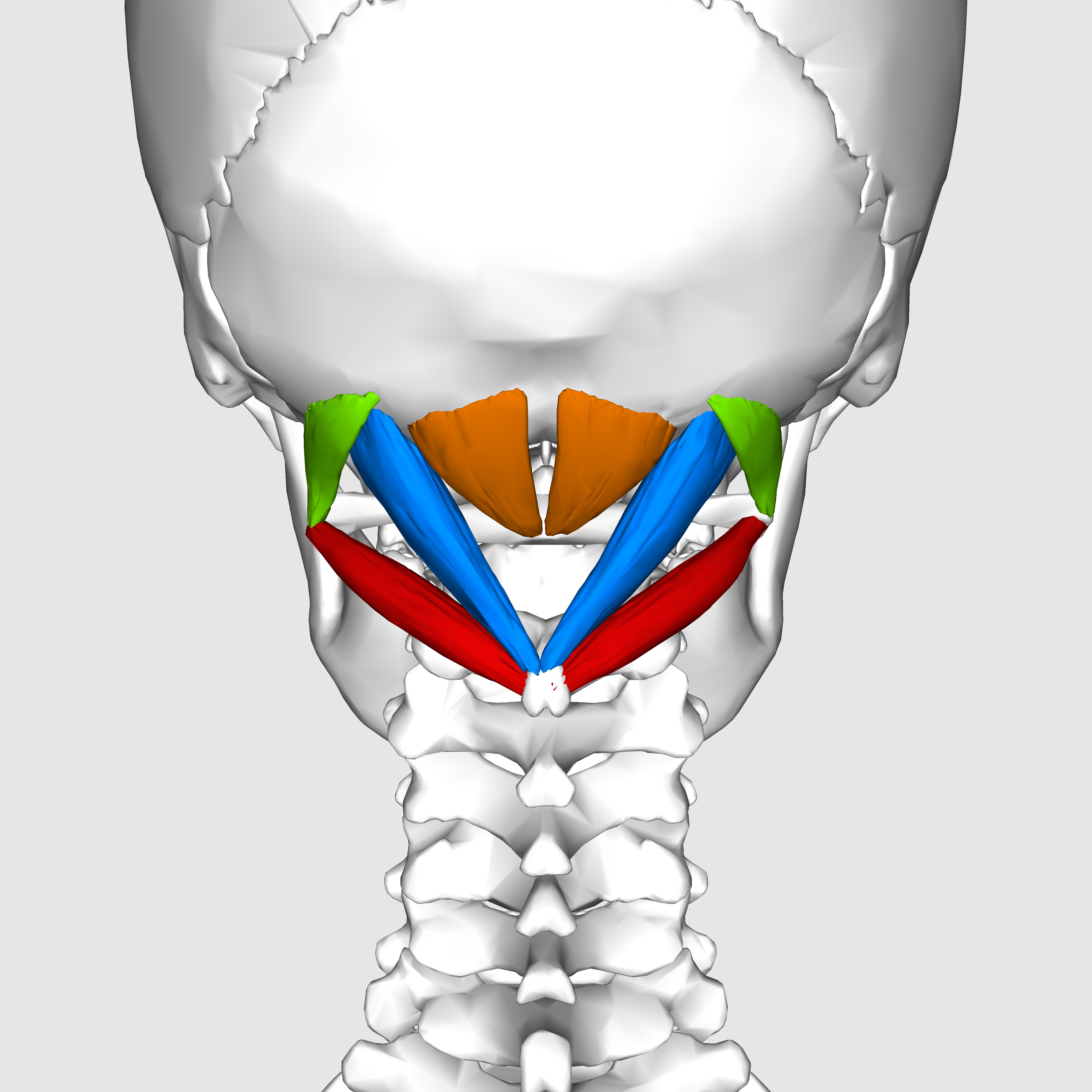
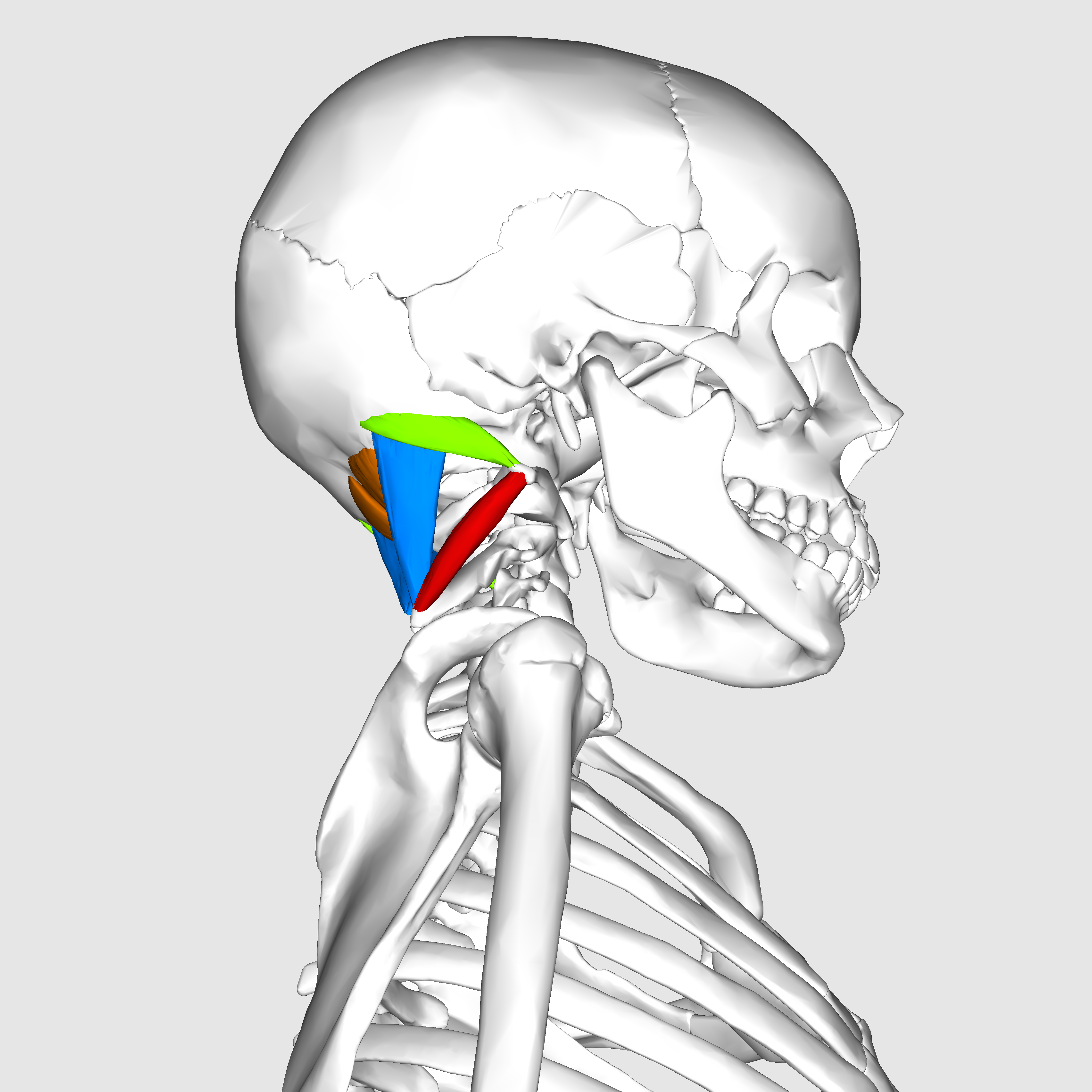
The suboccipital muscles connect the skull, the atlas vertebra, and the axis vertebra.
|
|---|
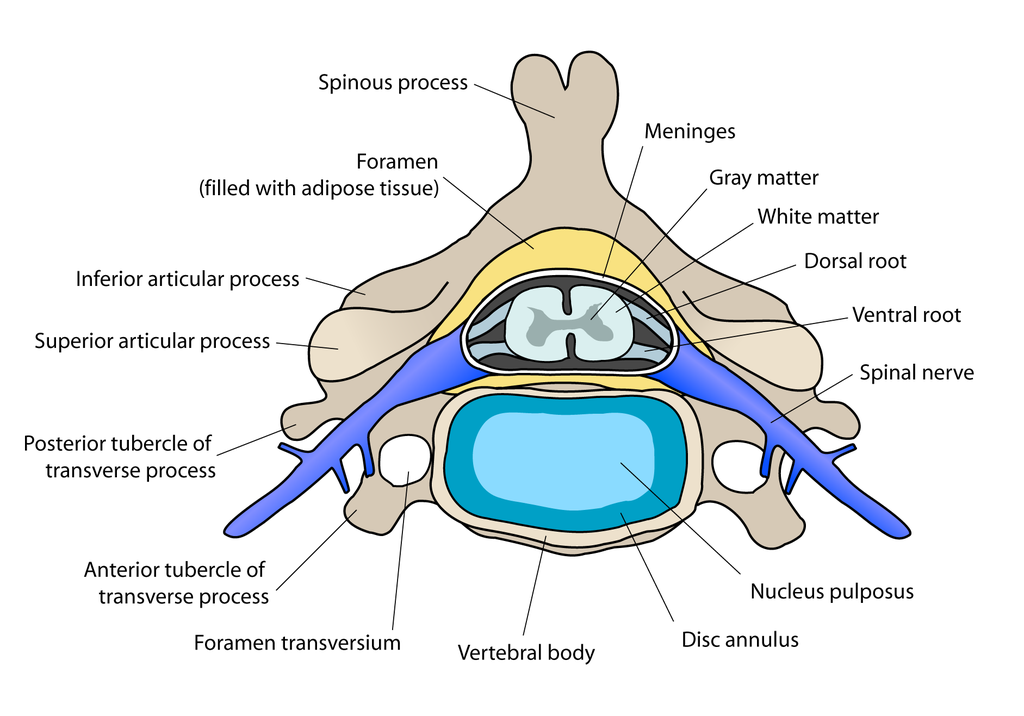
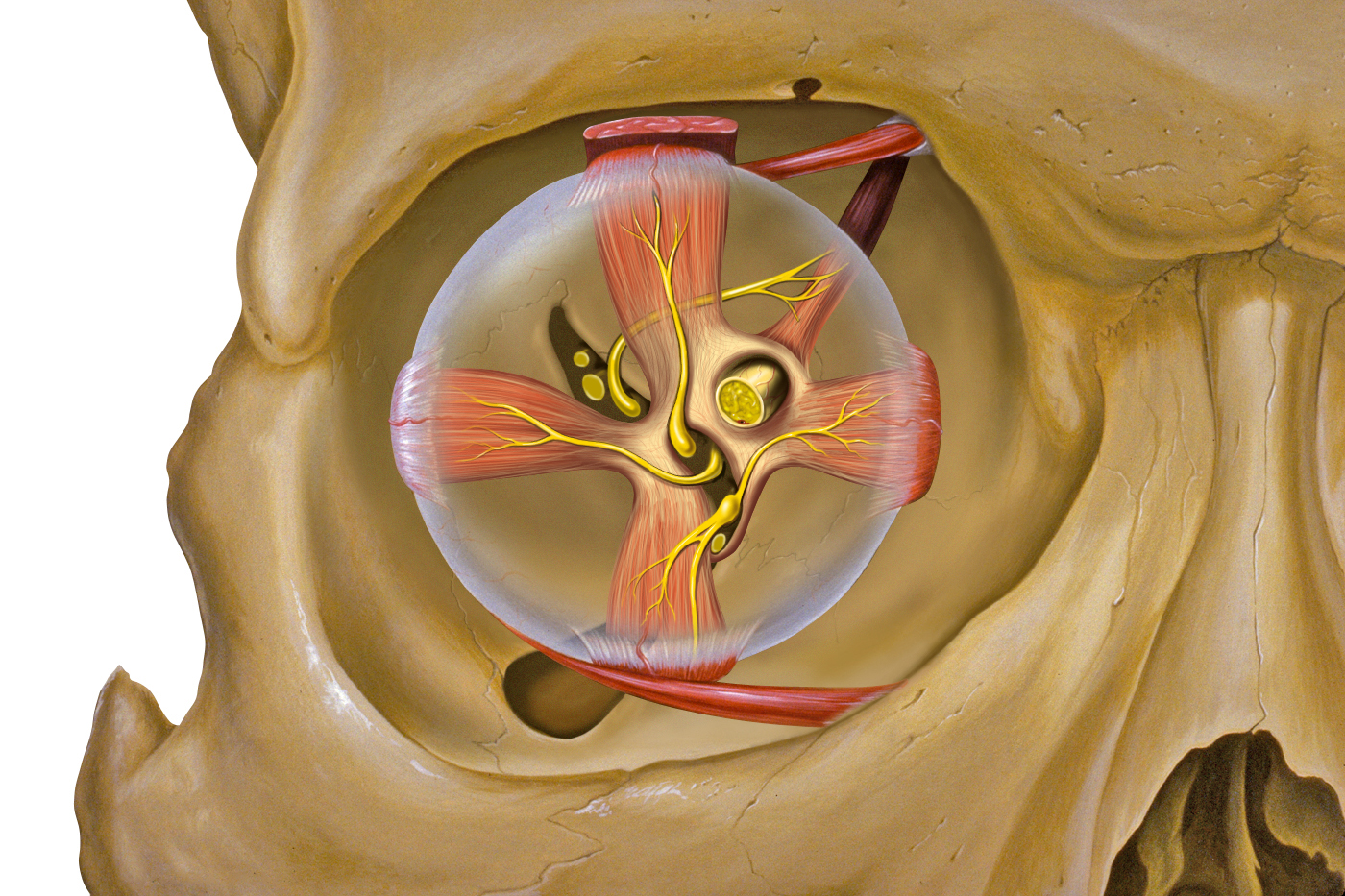
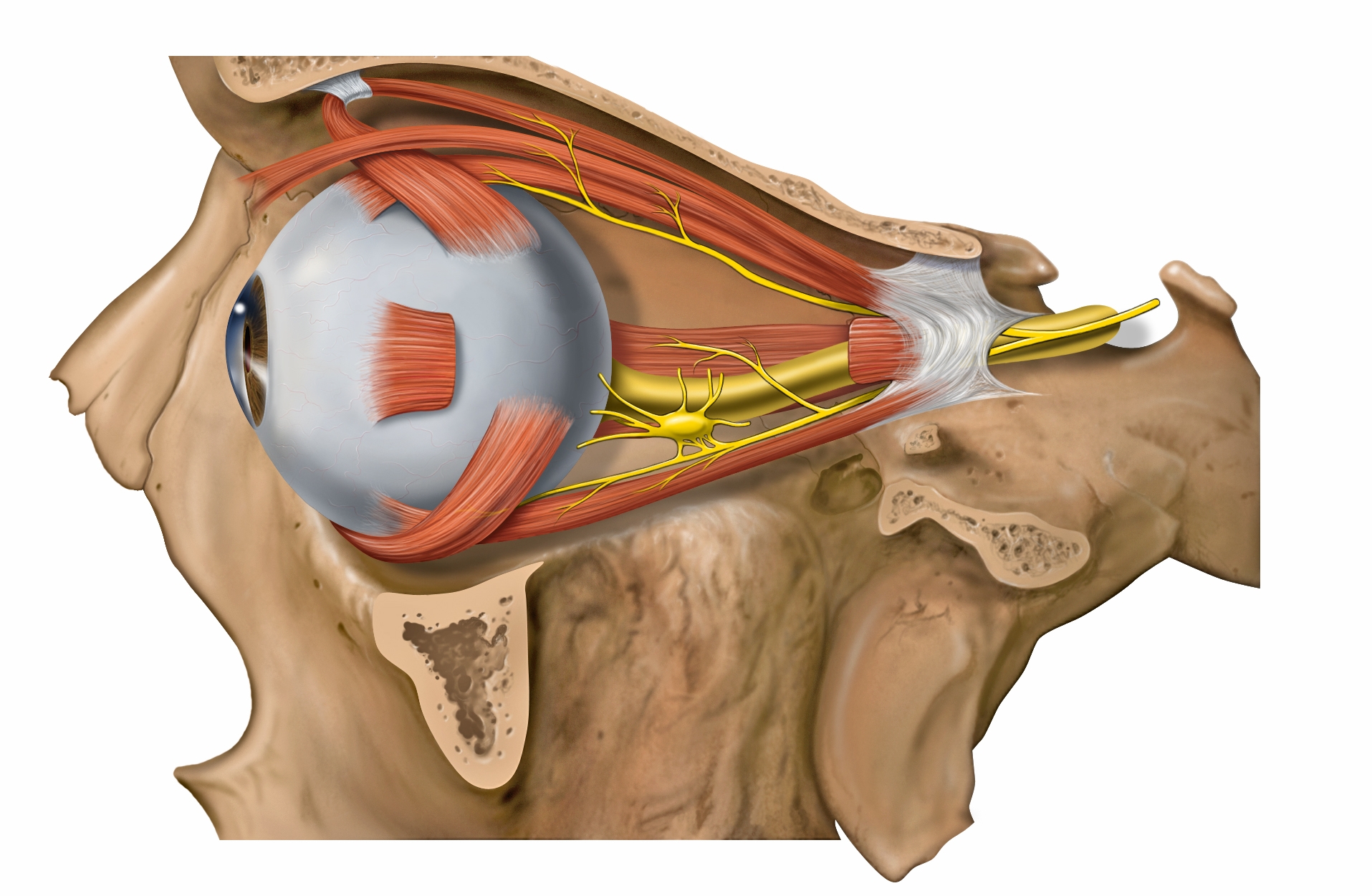
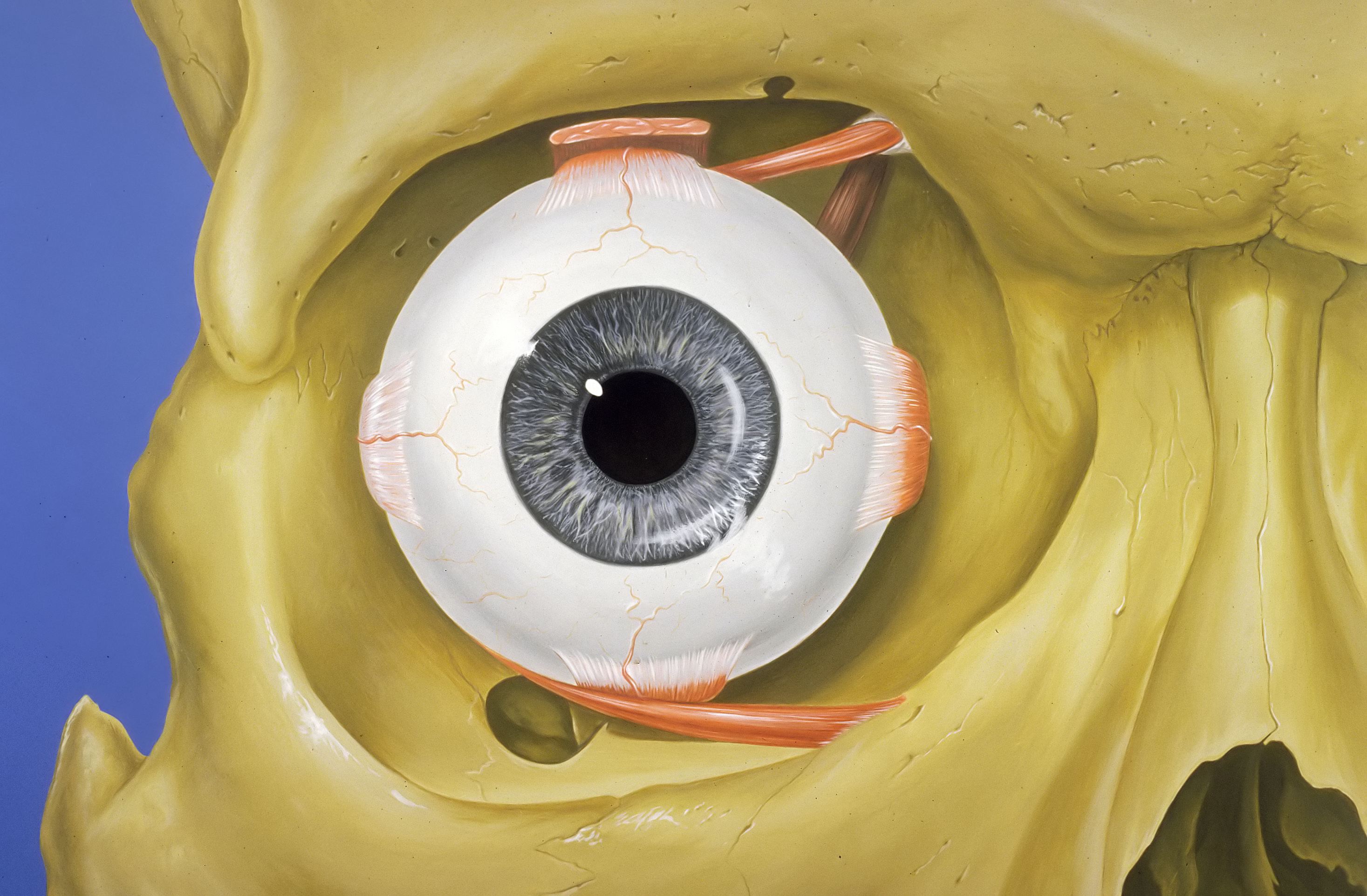
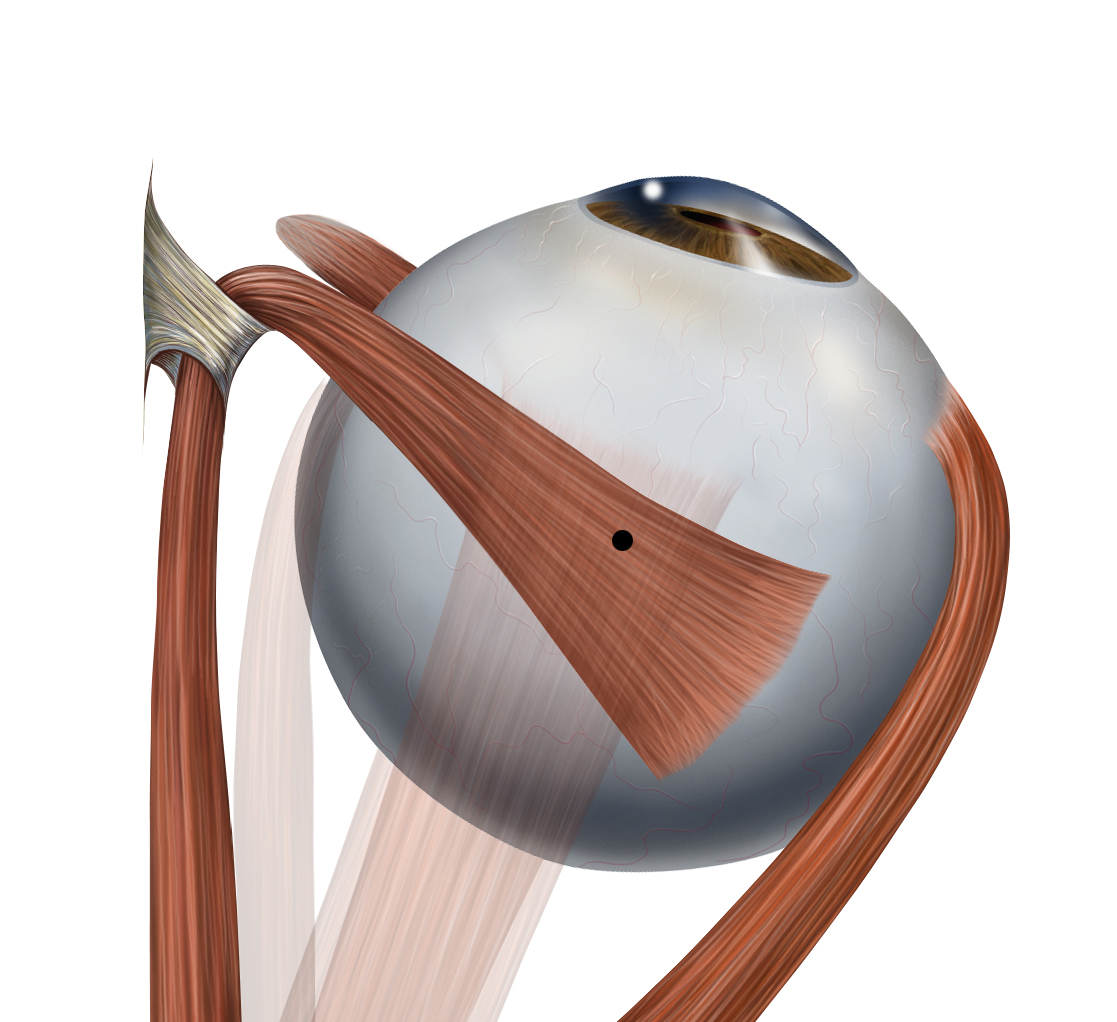
The atlas vertebra is at the center of the head and your eyes and ears are at the same level as the atlas vertebra.
The center of mass of the skull is slightly forward of the contact point between the skull and the atlas vertebra. If the muscles in your neck relax then your head pitches forward. If your back muscles relax then your torso pitches forward. The muscles in the back of your neck and spine act reflexively to prevent you from falling forward. This motion is coordinated with the breathing cycle.
 |
 |
|---|---|
Bruce Lee: When the opponent expands I contract, when he contracts I expand, and when there is an opportunity, I do not hit, it hits all by itself.
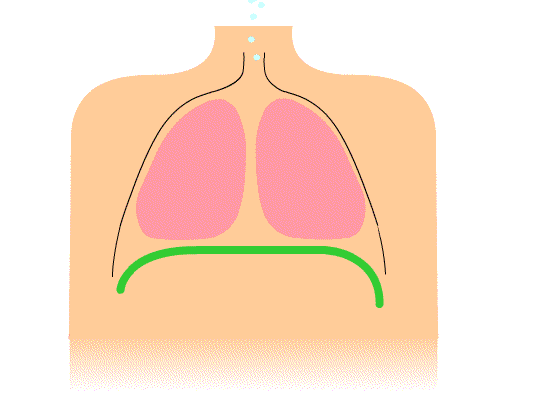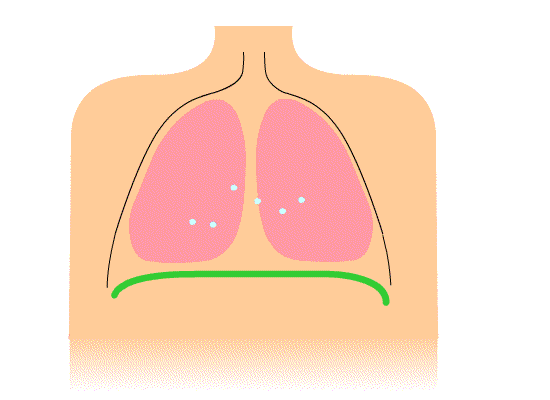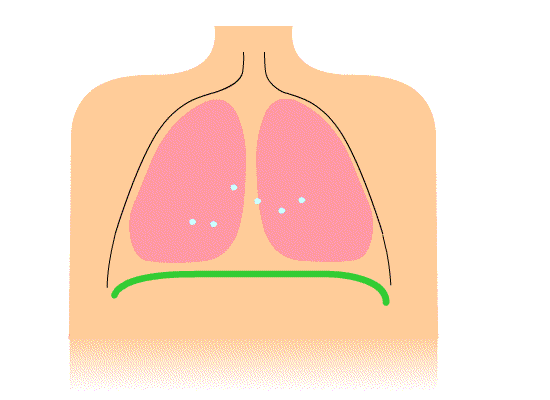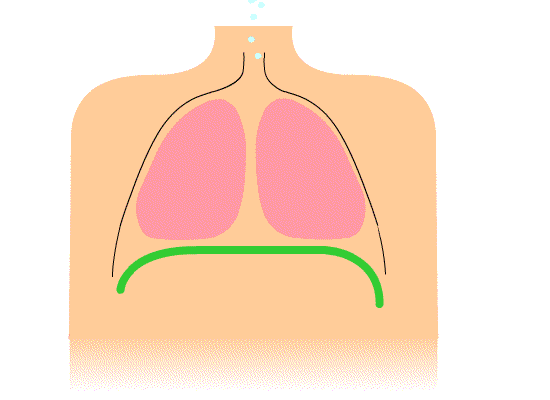
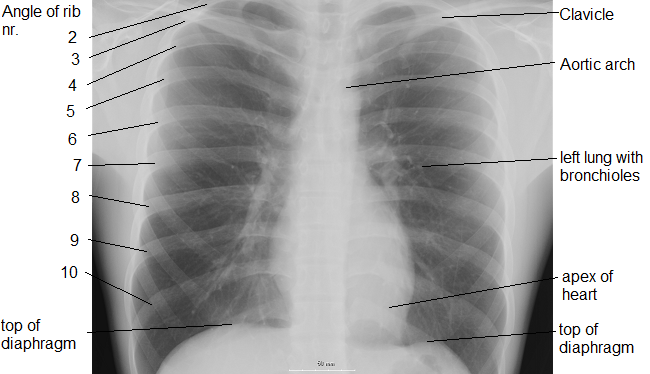
The thoracic diaphragm is underneath the lungs. When it contracts it creates overpressure in the gut, expands the ribcage, and creates underpressure in the lungs. The lung underpressure brings in air. Air is expelled from the lungs by contracting the rib intercostals.
 |
|---|
The pelvic diaphragm works in opposition to the thoracic diaphragm. When the thoracic diaphragm contracts the pelvic diaphragm expands and vice versa. This moves the gut cyclically up and down.
Breathing is coordinated with skeletal motion to minimize energy expense. For example, when you breathe in your head tends to pitch back and when you breathe out your head tends to pitch forward. It can be done the opposite way but it's less natural.
Every bilaterally symmetric motion is related to the breathing cycle according to the following table:
Inhale Exhale Diaphram contracts Diaphragm expands Abdominals expand Abdominals contract Gut squashed by diaphram Gut expands External intercostals Internal intercostals Ribcage expands Ribcage contracts Pelvic floor expands Pelvic floor contracts Spine muscles contract Spine muscles release Arms out Arms in Head pitches up Head pitches down Head rolls right Head rolls left Head yaws right head yaws left Arms rotate thumbs up Arms rotate thumbs down Elbows rotate out Elbows rotate in Open hand Form fist Feet rotate outward Feet rotate inward Knees apart Knees together Lower back arches Lower back sags Hips rotate forward Hips rotate back Daydream Focus Rebalance Exertion High moment of inertia Low moment of inertia Discard angular momentum Discard pressure Tongue makes "U" shape Tongue makes flat shape, such as for the letter "L" Spread fingers into a fan Bring fingers together like a fin
Rows correspond to opposite directions of a motion, and columns show how they synchronize with the breathing cycle.
Bilaterally antisymmetric motions are coordinated through the axis vertebra. Motion naturally cycles between the left and right columns in the table below:
Head rotates left Head rotates right Right hand rotates thumbs-up Right hand rotates thumbs-down Left hand rotates thumbs-down Left hand rotates thumbs-up Right foot rotates right Right foot rotates left Left foot rotates right Left foot rotates left Right arm rotates out Right arm rotates in Left arm rotates in Left arm rotates out Jaw pivots right Jaw pivots left Tongue pivots right Tongue pivots left Eyes pivot right Eyes pivot left Head rolls right Head rolls left Shoulders rotate right Shoulders rotate leftThe axis cycle is structured to most naturally conserve angular momentum. For example, if your right arm rotates thumbs-down then conservation of angular momentum is satisfied if your left arm rotates thumbs-up.
Cycle Nexus Head motion Motion type Goal Breathe Atlas vertebra Pitch Bilaterally symmetric Minimize energy expense Axis Axis vertebra Yaw Bilaterally antisymmetric Minimize internal angular momentum
When you move your head, it is most natural to coordinate rightward roll with rightward yaw, and leftward roll with leftward yaw. It can be done the opposite way but it's less natural.
The reason roll and yaw are related this way is because the center of mass of your head is forward of the balance point between your spine and skull. If you start from an upright position, roll right, and then stop, then conservation of angular momentum causes your head to yaw right.
The natural relationship between pitch and yaw is:
Upward pitch goes with righward yaw.
Downward pitch goes with leftward yaw.
Ideally, yaw should be minimized when the head pitches.
The prime balance foot is the right foot. If you stand on two feet your weight naturally shifts to the right foot. As your head moves to the right it yaws right (because the skull center of mass is forward of the spine). Moving to the right foot is also a descent in gravitational energy, which is countered by your head pitching back.
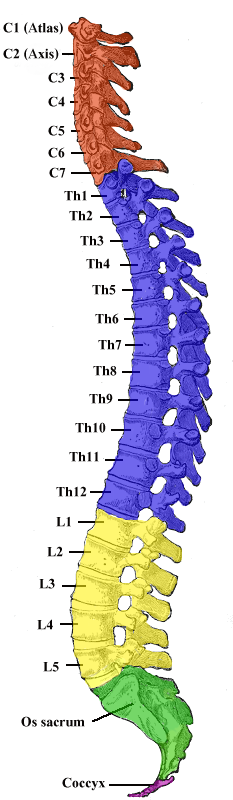
The spine consists of a set of curves that function as shock absorbers and as vertical motion for the head.
 |
 |
|---|---|
 |
 |
|---|---|
 |
 |
 |
|---|---|---|
 |
 |
|---|---|
 |
 |
 |
|---|---|---|
 |
 |
|---|---|
 |
 |
 |
|---|---|---|
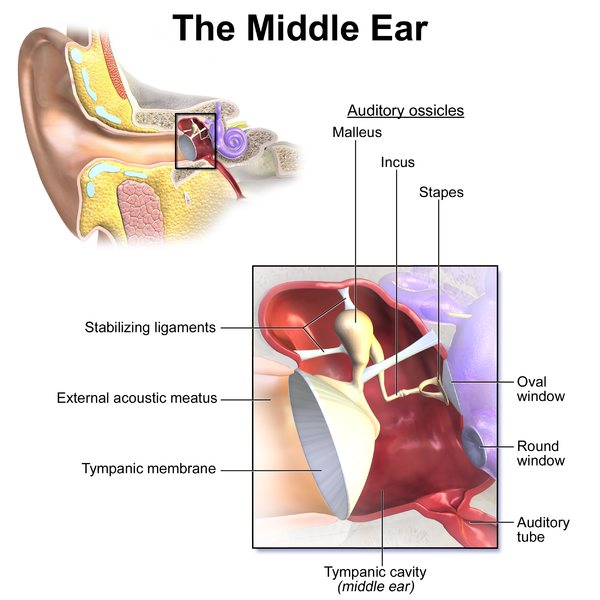
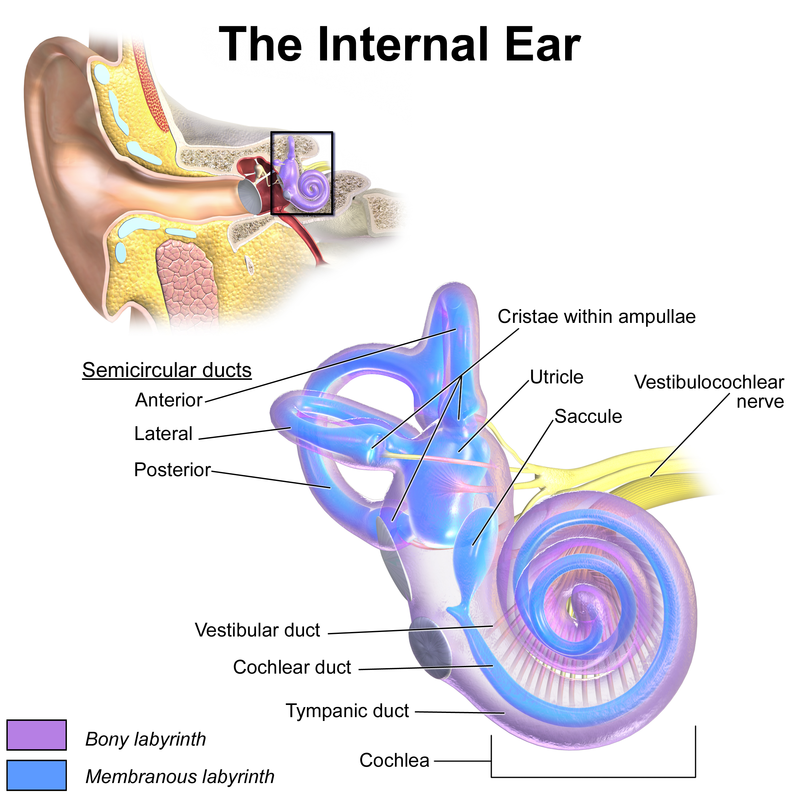
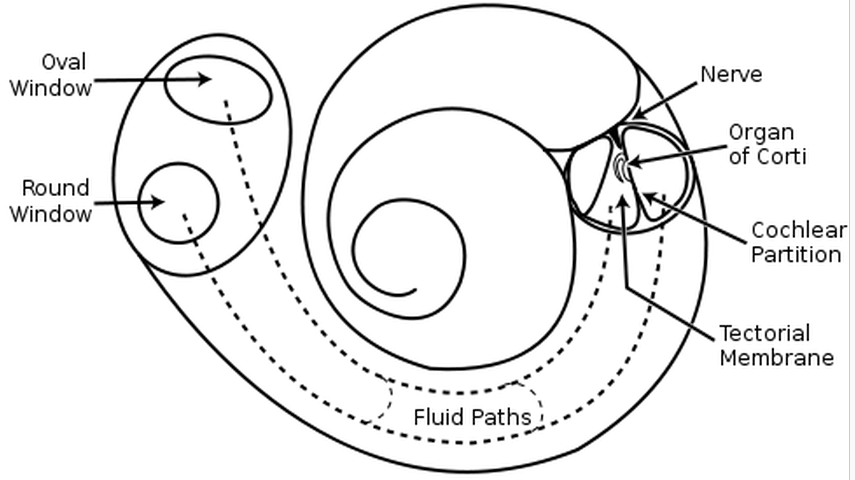
The timpanic membrane converts air pressure waves into mechanical motion of the ear bones. The ear bones amplify the signal and transmit it to the stapes bone, which is connected to the oval window of the cochlea. Vibrations in the oval window cause vibrations in the fluid of the cochlea, where they are converted into neural signals and interpreted in the brain.
If a sound wave in air encounters water then 1/30 of the sound energy is transmitted to the water and the rest reflects back into the air. If sound waves were transmitted directly from the air to the fluid of the cochlea then they would suffer this loss. The ear bones function to improve the transmission efficiency from the air to the fluid of the cochlea.
The tympanic membrane has 13 times the area of the oval window, and so the signal is amplified by a factor of around 13.
As pressure waves travel along the cochlea the cochlea narrows. The narrower the cochlea, the higher the frequency range it is sensitive to. Low frequencies are detected at the beginning of the cochlea and high frequencies are detected at the end.
If the sound level is too loud then the muscles of the middle ear shut down the motion of the ear bones. This is the "acoustic reflex" (Wikipedia).
The function of the ear bones was first explained by Helmholtz.
 |

|
|---|---|
A microphone records sound pressure as a function of time and a seismometer records displacement as a function of time. Your ears don't work anything like this. Your ears function instead detect the frequency spectrum, analogous to a spectrum of light.
 |
|---|
 |
|
|---|---|
There are 20000 hairs arranged along the length of the cochlea, each tuned to a different frequency. Each hair functions as a resonator.
High frequencies are detected at the start of the cochlea and low frequencies are detected
The perceived loudness depends on the duration of the note. For notes less than .2 seconds the loudness is proportional to duration and for notes more than .2 seconds the loudness is independent of duration. This suggests that the cochlea functions like a resonator, because it takes time for a resonance to activate.
If the duration of a note is much longer than 1 second then our attention fades and the note seems less loud.
Our ability to resolve frequency depends on the sharpness of the resonators in the cochlea. The brain provides active feedback to sharpen the resonance and suppress resonances at nearby frequencies.
If there are two sounds with different frequencies, then if the frequencies are too close to each other they will interfere with each other in the cochlea, and if they are far enough apart they can be sensed independently. The frequency width for interference is on the order of a perfect fifth.
Noise tend to obstruct our ability to resolve pitch.
Nerve signals travel both from the cochlea to the brain and from the brain to the cochlea. The brain provides active feedback to refine the function of the cochlea.
There are nerves that travel directly back and forth between your ears for stereo processing.
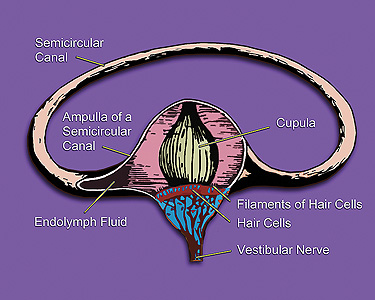
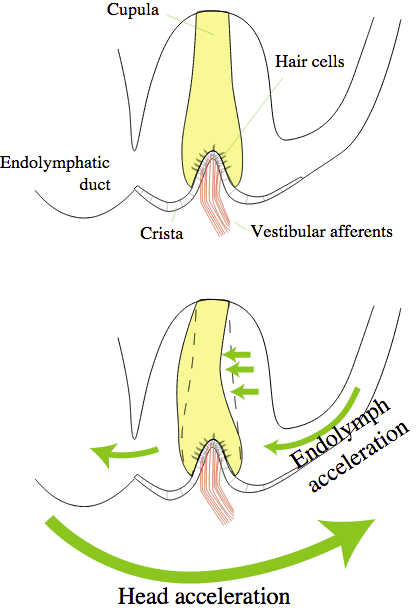
The semicircular canals of the cochlea are a gyroscope. Rotating your head causes fluid to flow in the canals, which is detected by hair cells. The function of the gyroscope and the function of the auditory system are connected.
The vestibule and the saccule are hardened objects used to detect linear acceleration. WHen you accelerate these objects are displaced, which is detected by hair cells.
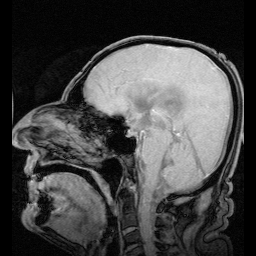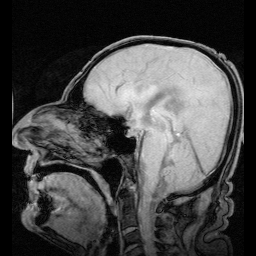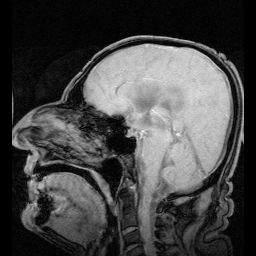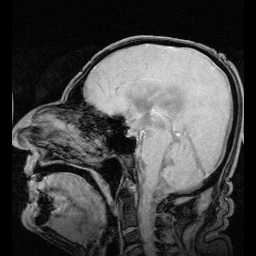
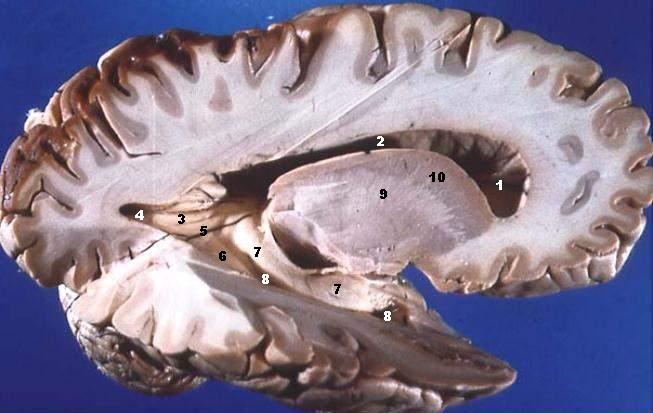
Your ears are at the center of your skull, aligned with the pivot point that connects your skull to the top of your spine. The ear is involved in calculating balance.
 |

|
|---|---|
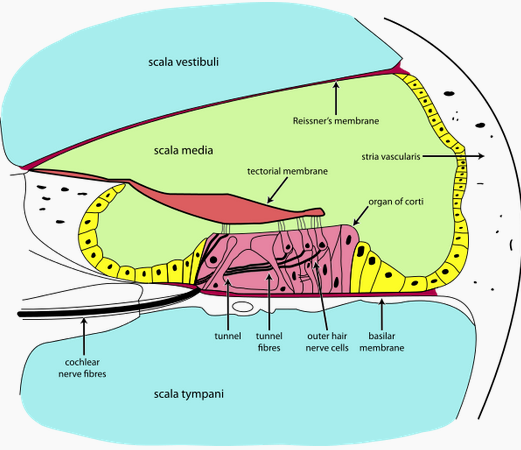
The basilar membrane functions like a harp or piano. It is a strip running the length of the cochlea, narrow at the end closest to the ear and wide at the end farthest from the ear, like a necktie. It is also stiffer the closer it is to the narrow end. The resonant frequency at any particular point along the basilar membrane increases with stiffness and decreases with width, giving it a frequency range that varies from high to low as you traverse from the narrow to the wide end. Siffness is controlled with muscle tension.
The lower the frequency of the wave, the further it propagates along the basilar membrane. High-frequency waves diminish before they get to the wide end.
The fact that low frequencies propagate further along the basilar membrane is analogous to the fact that low-frequency pitches more easily pass through walls than high-frequency pitches. A low-frequency pitch has more time to move the wall for a given sound pressure.
Helmholtz was the first to characterize the function of the basilar membrane
One out of every 10000 people has "absolute pitch", where for example you can tell if a pitch is higher, lower, or equal to 440 Hertz. Everone else has "relative pitch", where pitch ratios can be sensed but not absolute pitch. This suggests that there is no fixed place on the basilar membrane that corresponds to 440 Hertz.
If you don't have absolute pitch it is difficult to acquire it. From Wikipedia: "There are no reported cases of an adult obtaining absolute pitch ability through musical training; adults who possess relative pitch, but who do not already have absolute pitch, can learn pseudo-absolute pitch, and become able to identify notes in a way that superficially resembles absolute pitch. Moreover, training pseudo-absolute pitch requires considerable motivation, time, and effort, and learning is not retained without constant practice and reinforcement."
The ear is sensitive to both pitch and time. Pitch is measured by position along the basilar membrane and time is measured by differences beween neural signals. For high-frequency pitches we are more sensitive to frequency and for low-frequency pitches we are more sensitive to time.
 |
 |
|---|---|
The pinna is the outer part of the ear that collects sound and helps in determing its directionality. All human pinna are unique in shape and if the shape were to change it would affect your ability to determine the direction of sound.
A large pinna can amplify sound by 10 to 15 decibels.
Suppose a sound pulse arrives at your ear and an echo arrives a time T later. If T < 30 ms then you don't notice the echo and if T > 30 ms you notice the echo.
The distance a sound wave travels in a time of 30 ms is 10 meters. A concert hall has to be smaller than this to not sound like it has echos.
To do echolocation you have to train your ears to be sensitive to intervals less than 30 ms.
Bats use high frequencies for echolocation because they diffract less than low frequencies and hence give better resolution.
 |
|---|
Anguy Keep your elbow high. You want your back doing the hard labor.
You're holding. Never hold.
Arya What?
Anguy Your muscles tense up when you hold. Pull the string back to the center
of your chin and release. Never hold.
Arya But I have to aim.
Anguy Never aim.
Arya Never aim?
Anguy Your eye knows where it wants the arrow to go. Trust your eye.
Bruce Lee: Experiments indicate that auditory cues, when occurring close to the athlete, are responded to more quickly than visual ones. Make use of auditory clues together with visual clues, if possible. Remember, however, the focus of attention on general movement produces faster action than focus on hearing or seeing the cue.
Bruce Lee: You hear the bird chirping? If you don't hear the bird you cannot hear your opponent.
 |
|---|
Meters/second
Neuron 100
Sound in air 343 At 20 Celsius
Sound in water 1482
Light 300000000
Time in milliseconds:
.000003 Time for light to cross a 10 meter orchestra
.2 Electric synapse. These synapses are 2-way and they do not amplify signals
.7 Time for water pressure wave to travel 1 meter through your body
2 Chemical synapse. These synapses are 1-way and they can amplify signals
1 Time for a neural signal to travel 10 cm, the size of a brain
10 Time for a neural signal to travel from your fingers to your brain
3 Time for sound to travel 1 meter, the distance to an adjacent musician
7 Period of a 130 Hertz wave. This is the frequency of a viola C string
30 Time for sound to travel across a 10 meter orchestra.
62 Time between notes in "Flight of the Bumblebee"
For an orchestra to have good timing it must use visual cues. Sound isn't fast enough.
This is especially true at the rear of the viola section amidst the cacophony
of winds and brass.
The Europa Galante uses precise visual cues.
Pressure waves in your muscles deliver information 15 times faster than neurons.
When listening to an orchestra one's attention most easily falls on the high-frequency instruments. Practice listening to the low-frequency instruments, especially the cellos and basses. They control the long-term temporal coherence.
Bruce Lee: You hear the bird chirping? If you don't hear the bird you cannot hear your opponent.
If you can't hear the violas you can't hear the chord. Practice listening to the middle note of the chord.
One should also practicing listening to instruments at minimal volume. Loud volume obstructs our ability to resolve pitch.
The lower the sound intensity, the more sensitive we are to pitch. Practice listening to music at minimal volume.
Anticipate the pitch in your mind before you play it. The cochlea has active feedback from the brain and this helps harness it.
Develop fast reactions to adjust the pitch to be in tune with the rest of the orchestra.
Frequency Wavelength (Hertz) (meters) 20 15 Lower limit of human frequency sensitivity 41 8.3 Lowest-frequency string on a string bass or bass guitar 65 2.52 Lowest-frequency string on a cello 131 2.52 Lowest-frequency string on a viola 440 .75 The A-string on a violin 660 .75 The E-string on a violin (highest-frequency string) 20000 .016 Upper limit of human hearing
 |
 |
|---|---|
 |
 |
 |
 |
|---|---|---|---|
 |
 |
 |
 |
|---|---|---|---|
The semicircular canals are .8 mm in diameter.
Youtube: Eye saccades in slow motion
The motor cortex is in front of the somatic cortex.
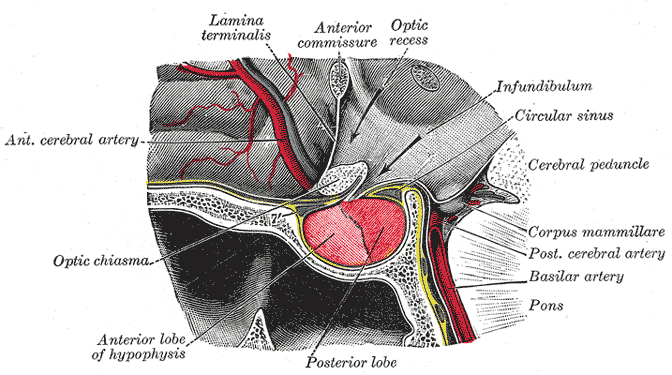
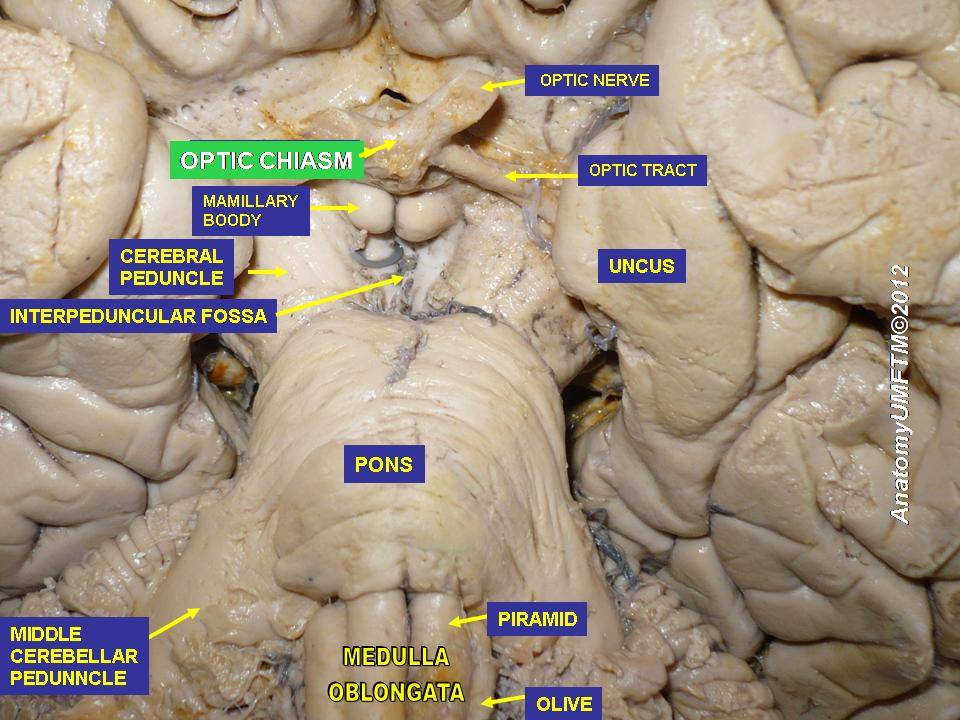
Information from the eyes crosses at the optic chiasm.
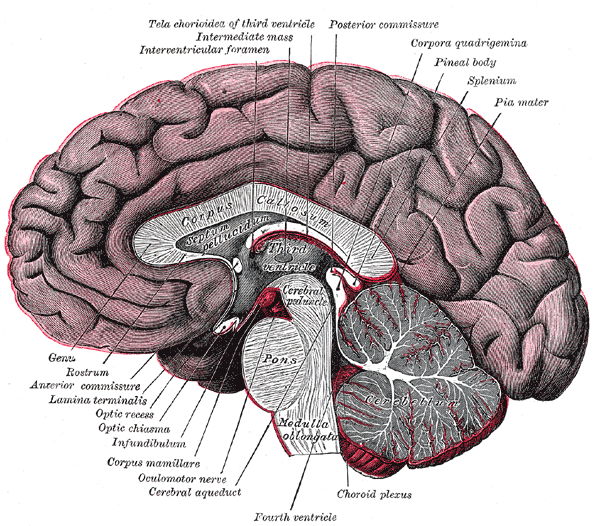
The corpus callosum connects the two brain hemispheres. It is tangibly larger
and more plastic in musicians.
Information from the eyes passes through the motor cortex before being
assembled at the rear of the brain. The motor cortex is an image-stabilization
system for the eyes. Visual input requires neural processing before it can be
interpreted.
In the motor cortex, proceeding from the center to the edge of the brain
corresponds to proceeding from the feet to the head of the body. It represents
a stack of reference frames starting from the ground and proceeding upward.
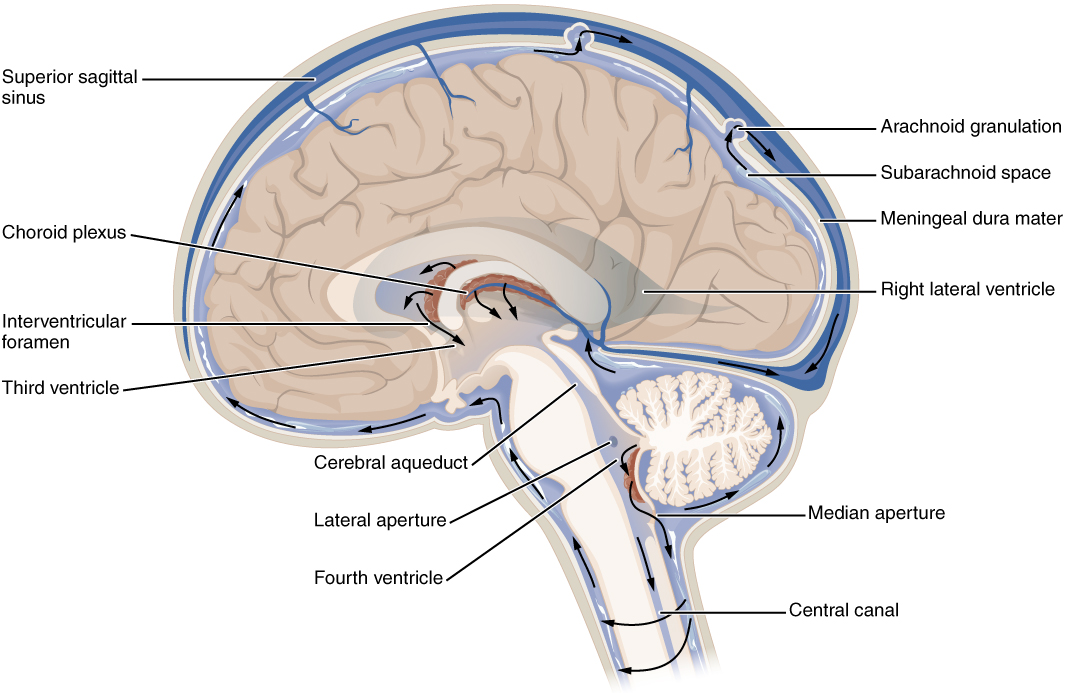
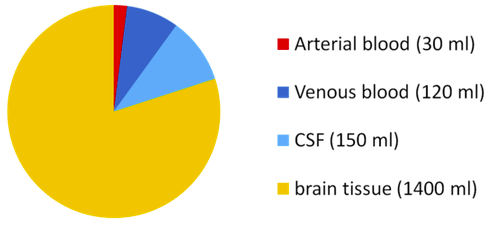
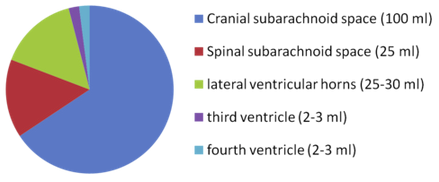
The brain produces 500 mL of cerebrospinal fluid per day and at any given time there
is 100-160 mL present.
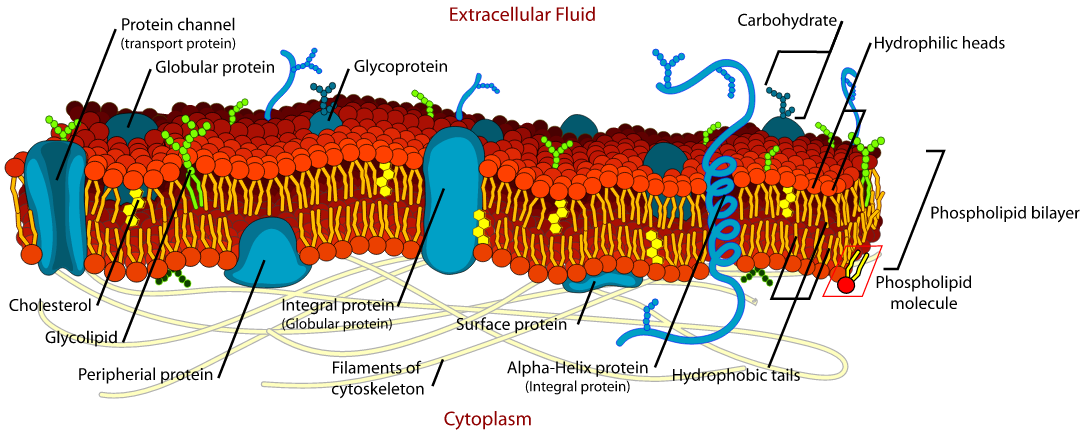
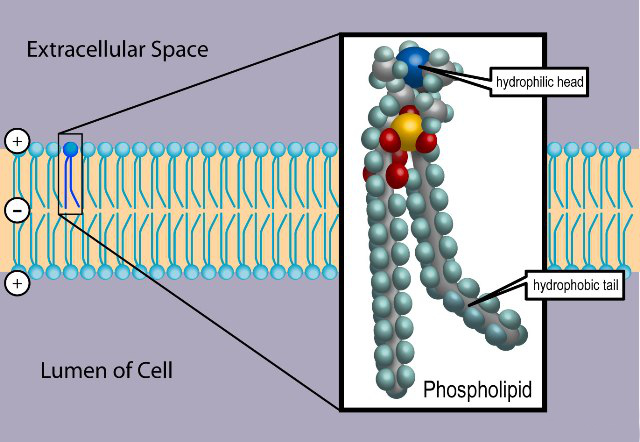
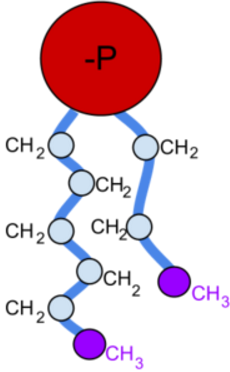
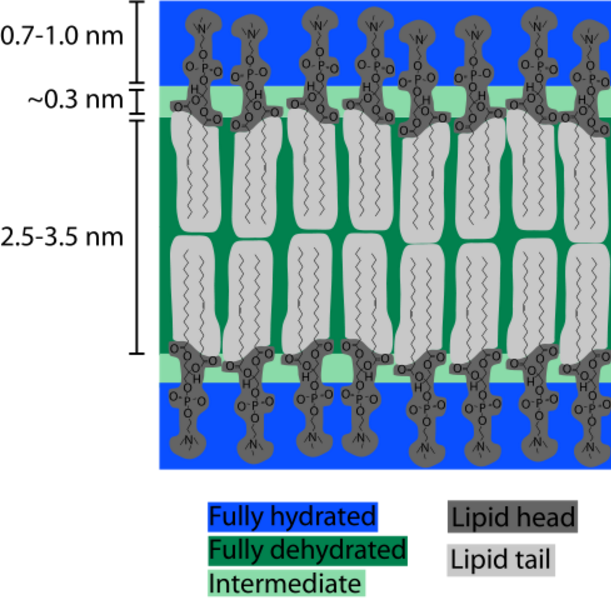
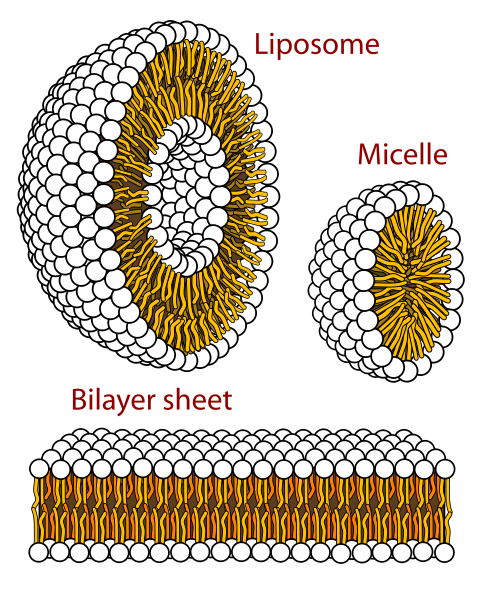
Cell membranes assemble spontaneously from phospholipid molecules. They are
mechanically flexibible due ot the ability of the phospholipids to rearrange
themselves.
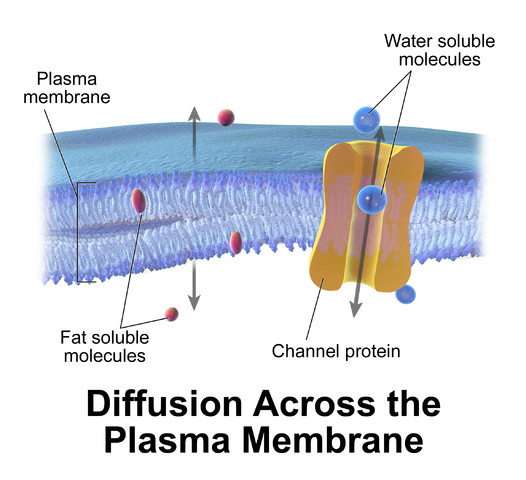
Cell membranes pass fat-soluble molecules and block water-soluble
molecules. Proteins can move molecules across the membrane.
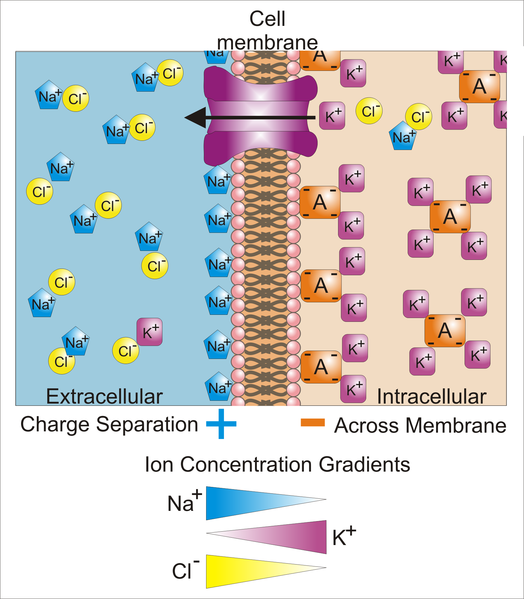
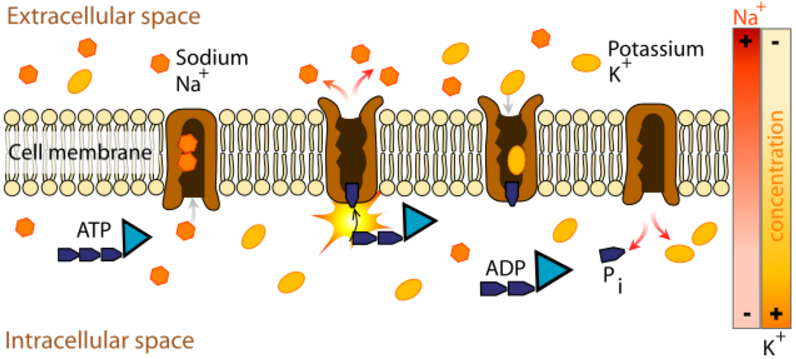
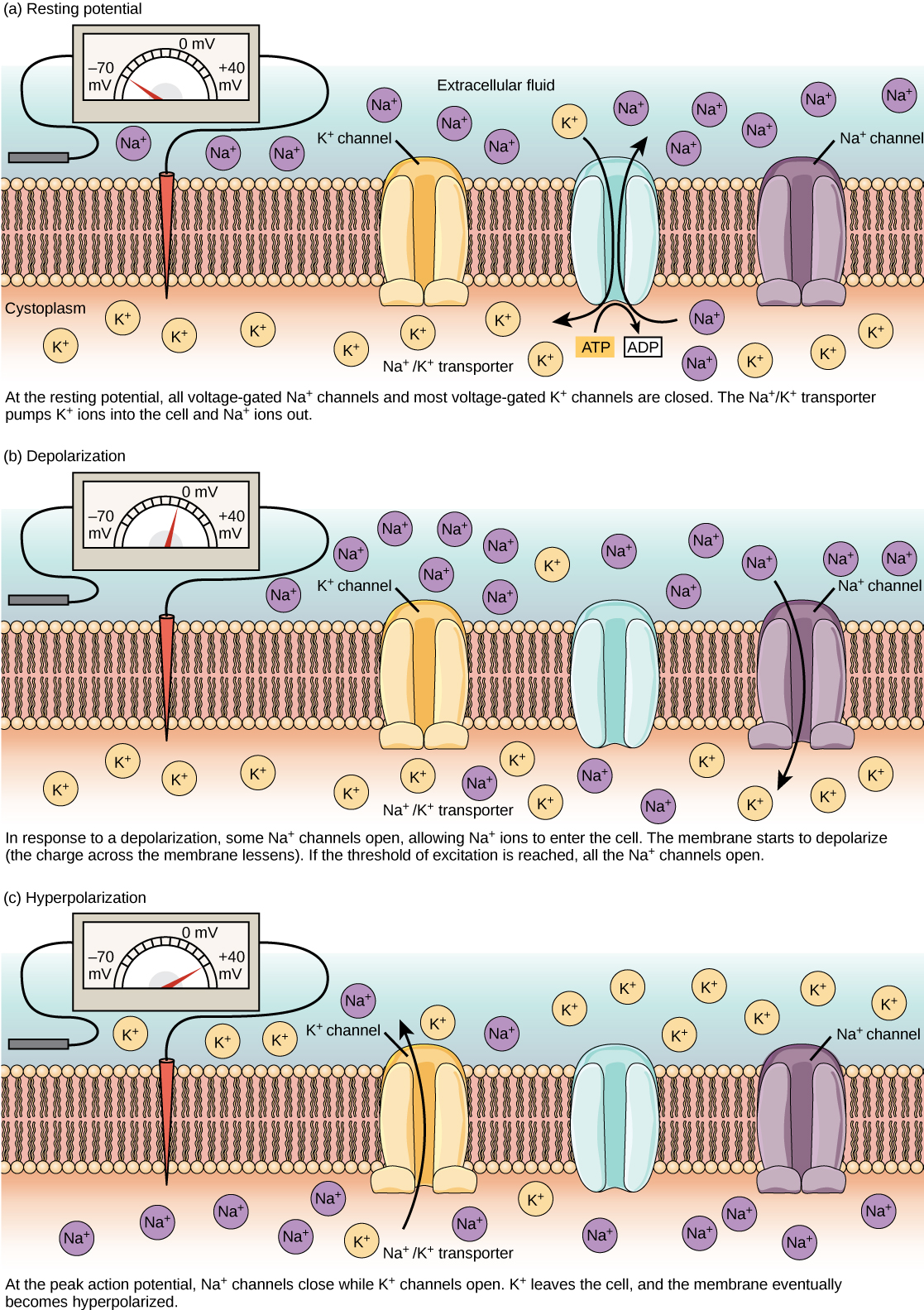
A membrane has ion channels that passively permit ion flow, and ion pumps that
actively transport ions. Most ion channels are permeable only to specific types
of ions. Ion channels can be modulated by either the membrane voltage or by chemicals.
The sodium-potassium pump generates a membrane voltage of around 70 mVolts, with
the cell interior being negative.
In each cycle of the sodium-potassium pump, 3 sodium ions move outward and 2
potassium ions move inward. The pump requires hours to establish equilibrium.
The pump is powered by ATP and the voltage gradient it produces provides a
power source for other ion pumps.
In each cycle of the sodium-calcium pump, 3 sodium ions move inward and
1 calcium ion moves outward. This pump is powered by the membrane potential
and doesn't require ATP.
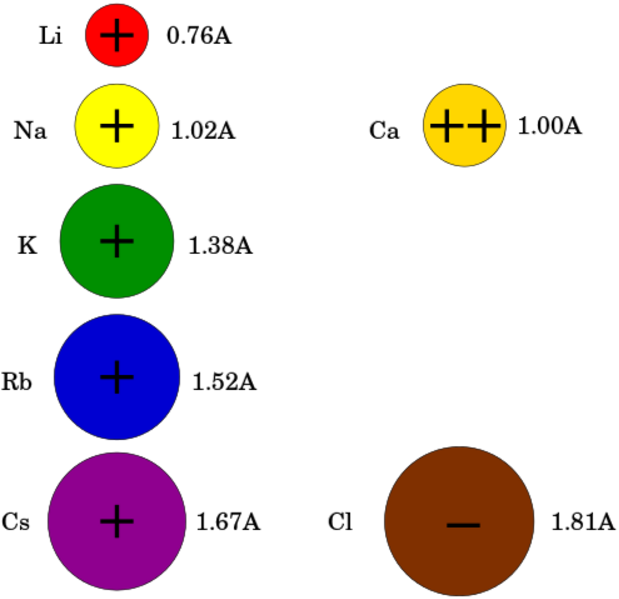
An ion channel for a given ion does not pass larger ions, and
most ion channels are specific for one ion. For example, most potassium
channels are characterized by 1000:1 selectivity ratio for potassium over
sodium, though potassium and sodium ions have the same charge and differ only
slightly in radius. The pore is small enough so that ions must pass
in single-file.
An action potential involves the opening and closing of ion channels and
doesn't involve ion pumps. If the ion pumps are turned off by removing their
energy source, or by adding an inhibitor such as ouabain, an axon can still
fire hundreds of thousands of action potentials before the amplitudes begin to
decay significantly.
The chloride ion is not actively pumped and takes on an equilibrium
concentration given by the membrane potential.
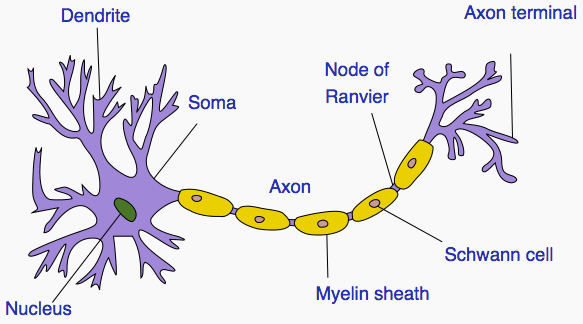
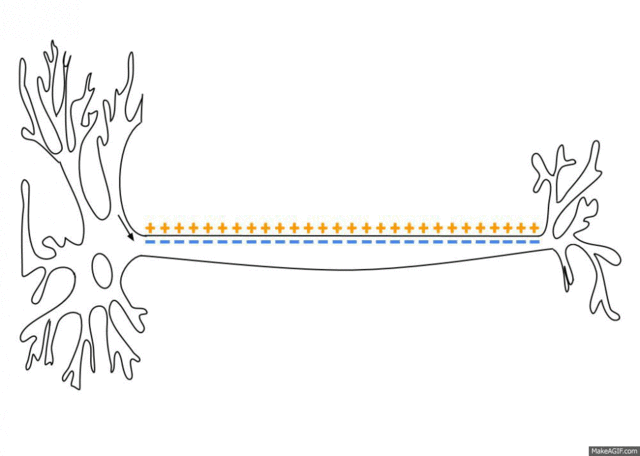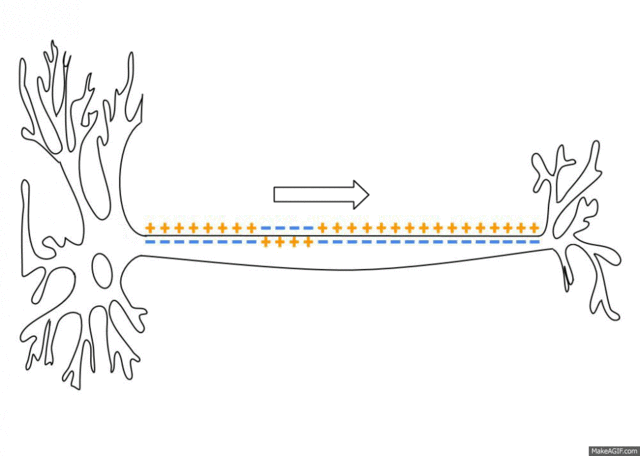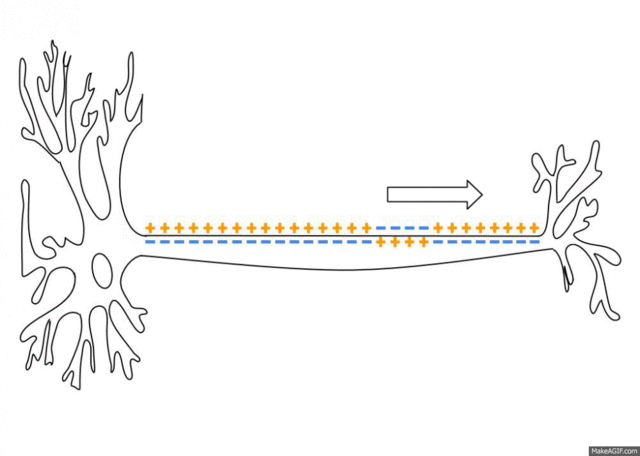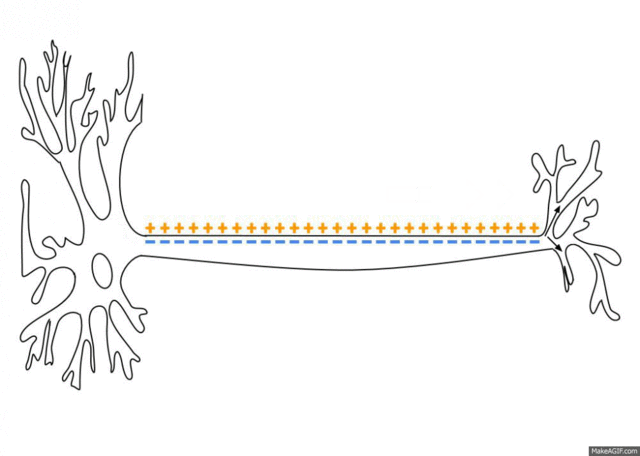
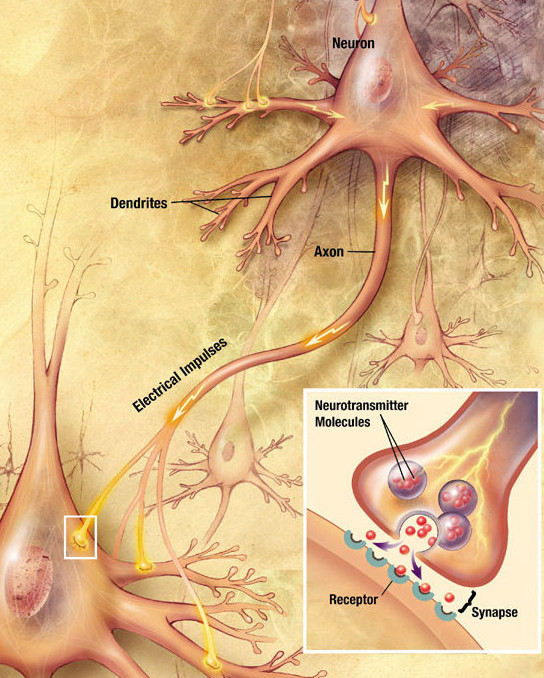
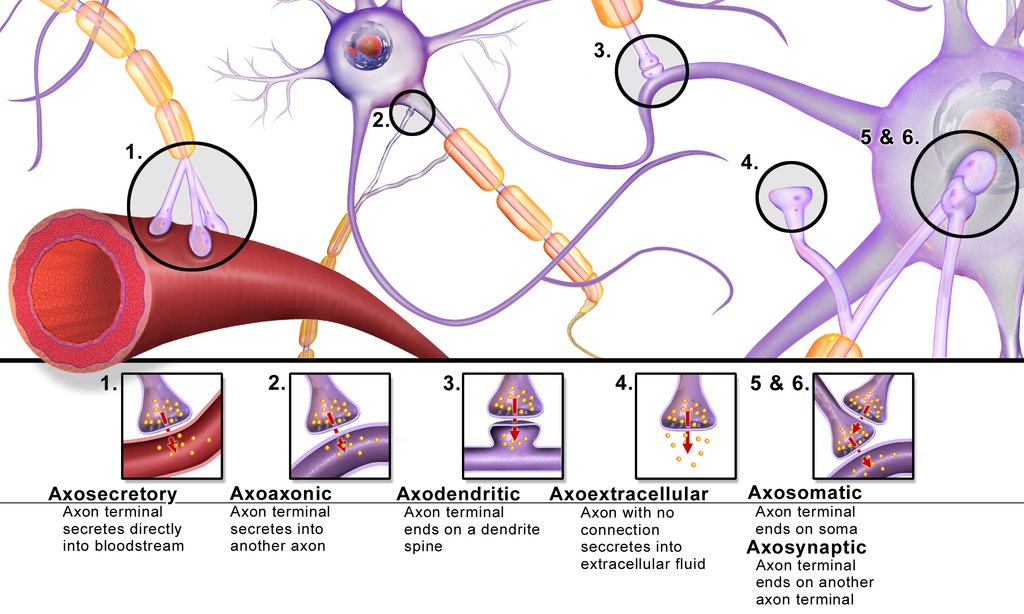
Signals travel from the cell body outward along an axon, jump to the dendrite
of another neuron at a synapse, then travel inward along the dendrite
to the cell body of the new neuron.
A neuron has at most one axon but the axon can branch hundreds of times. A
neuron can have multiple dendrites. There are, however, many exceptions to
these rules: neurons that lack dendrites, neurons that have no axon, synapses
that connect an axon to another axon or a dendrite to another dendrite, etc.
In certain sensory neurons (pseudounipolar neurons), such as those for touch
and warmth, the electrical impulse travels along an axon from the periphery to
the cell body, and from the cell body to the spinal cord along another branch
of the same axon.
The longest axons in the human body are those of the sciatic nerve, which run
from the base of the spinal cord to the big toe of each foot. The diameter of
axons is also variable. Most individual axons are microscopic in diameter
(typically about one micrometer across)
1) Unipolar neuron. Axon and dendrite emerging from same process.
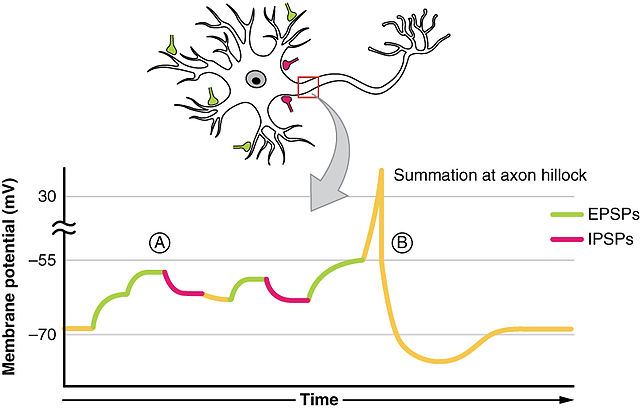
Axons connect to the cell body through the axon hillock. The axon hillock is
the last site in the cell where membrane potentials propagated from synaptic
inputs are summated before being transmitted to the axon.
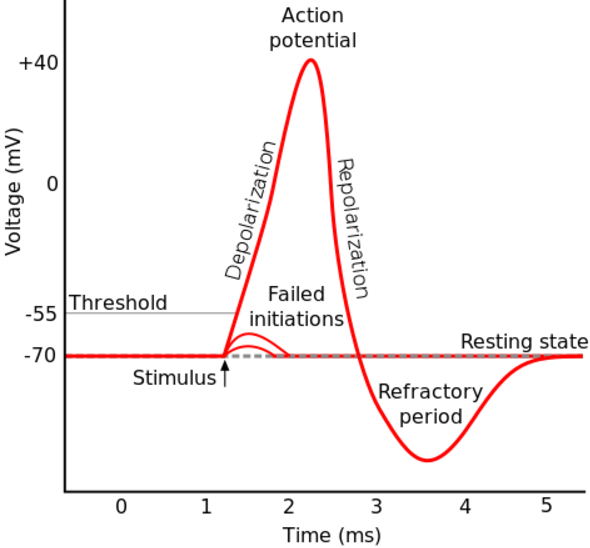
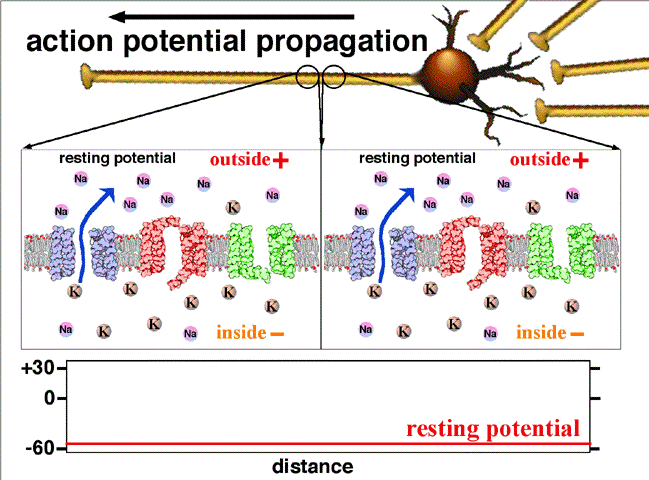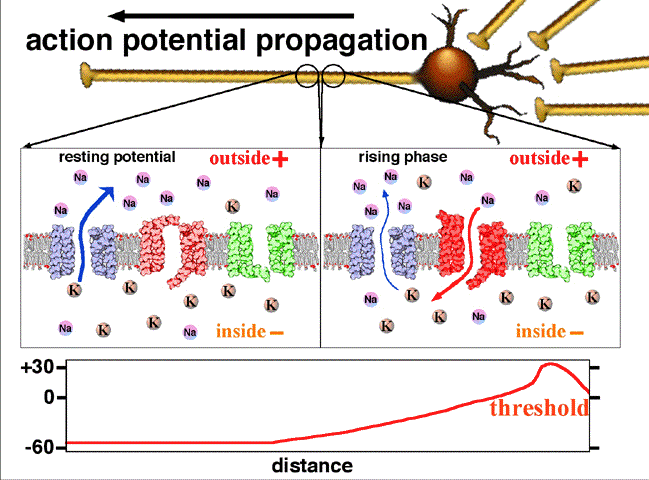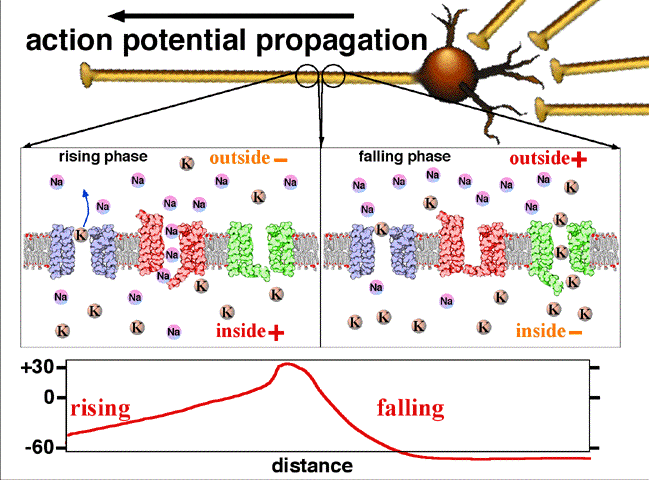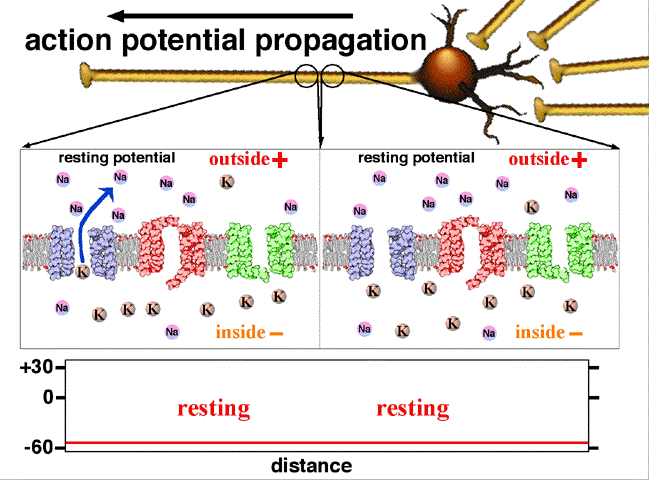
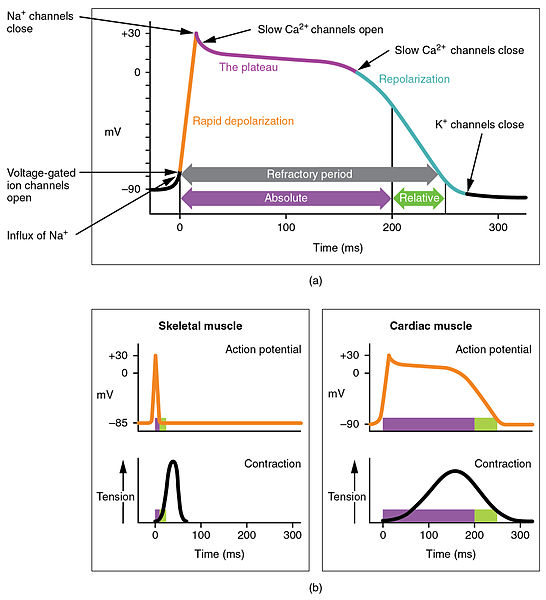
If the voltage across the membrane exceeds the threshold, voltage-gated sodium
ion channels open and sodium rushes into the cell, accelerating the voltage
rise. When the voltage reaches its peak the sodium channels close and potassium
channels open, restoring the potential to its resting state.
If the voltage change is too small to cross the threshold,
the potassium
current exceeds the sodium current and the voltage returns to its normal
resting value.
After the action potential fires the axon enters a refractory state, which
is responsible for the unidirectional
propagation of action potentials along axons. At any given moment, the
patch of axon behind the actively spiking part is refractory, but the patch in
front, not having been activated recently, is capable of being stimulated by
the depolarization from the action potential.
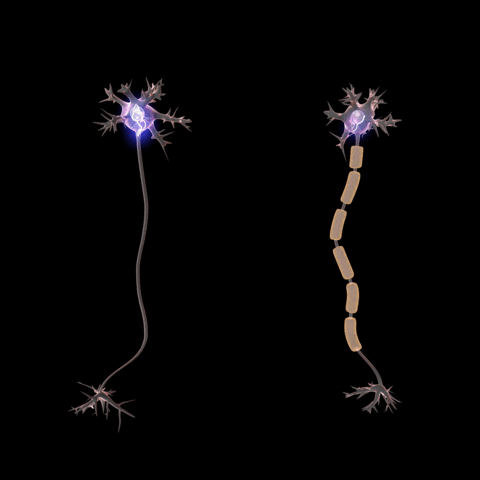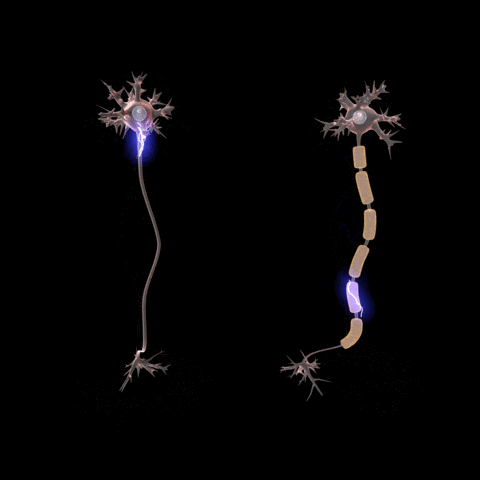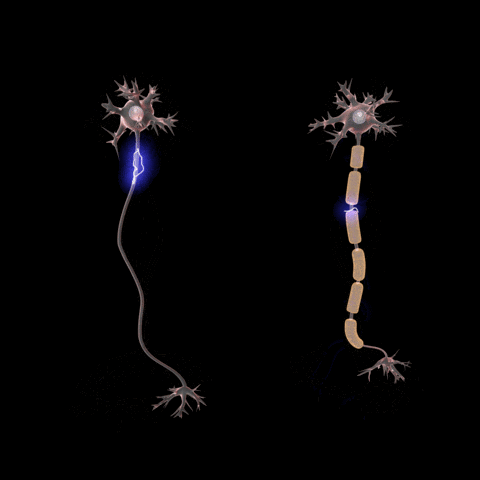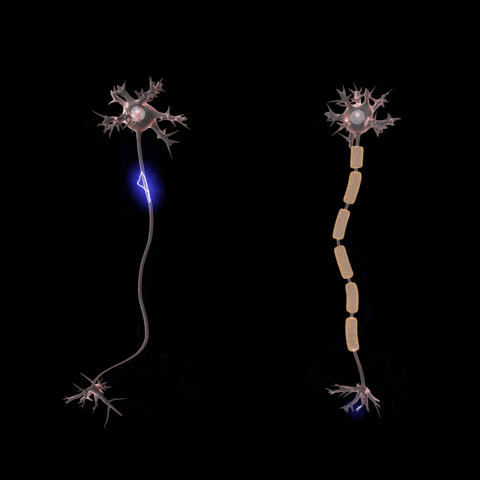
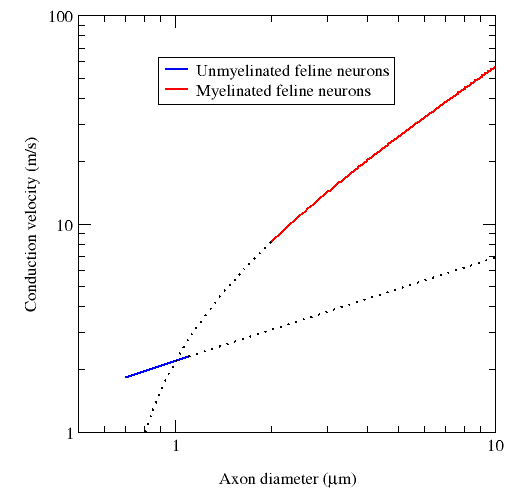
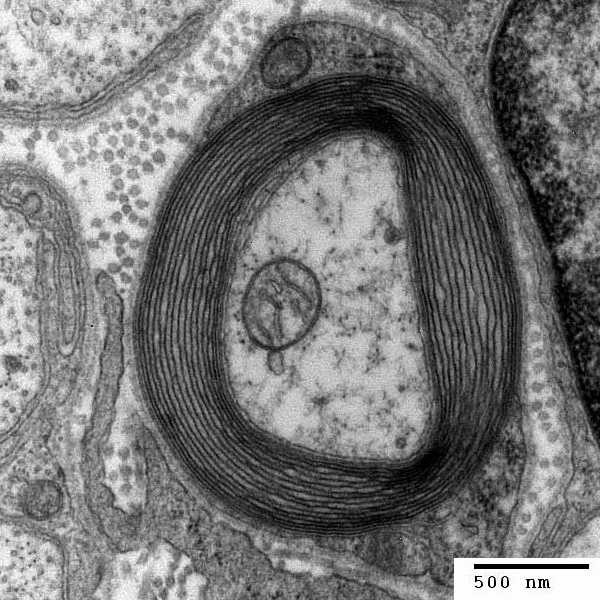
A myelin coating increases the speed of signals. Myelinated axons are known as
nerve fibers.
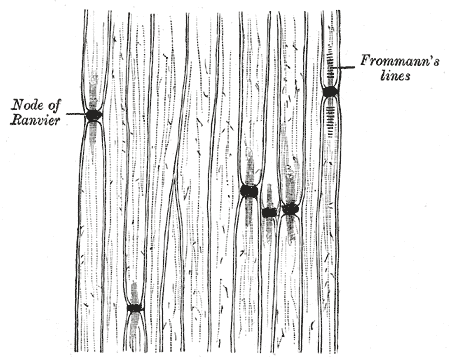
Signal propatation in myelinated axons is called "saltatory conduction", where
the signal jumps rapidly from one node of Ranvier to the next.
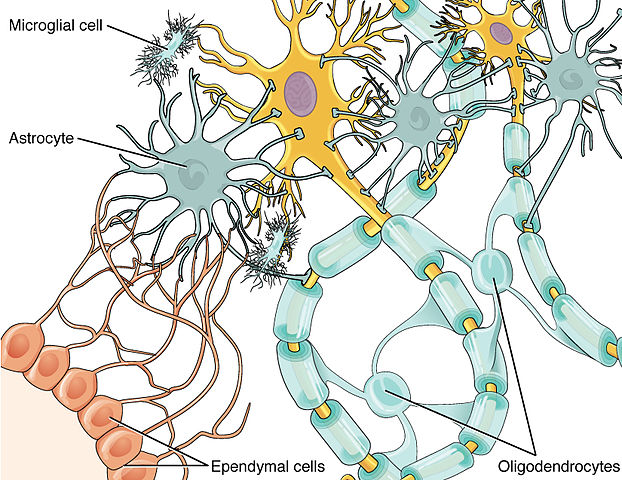
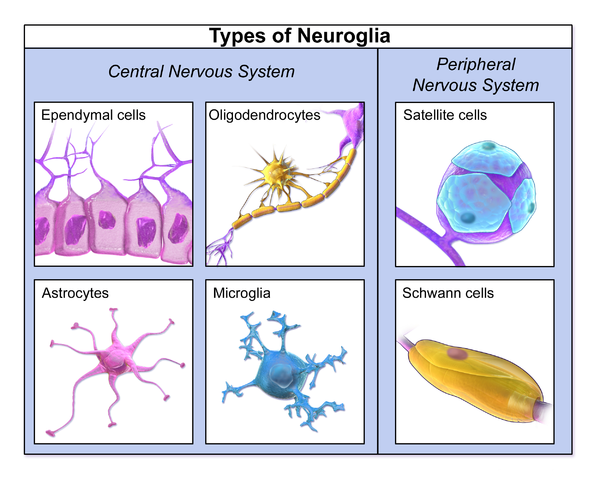
In the central nervous system (CNS), myelin is produced by oligodendroglia
cells. Schwann cells form myelin in the peripheral nervous system
(PNS). Schwann cells can also make a thin covering for an axon which does not
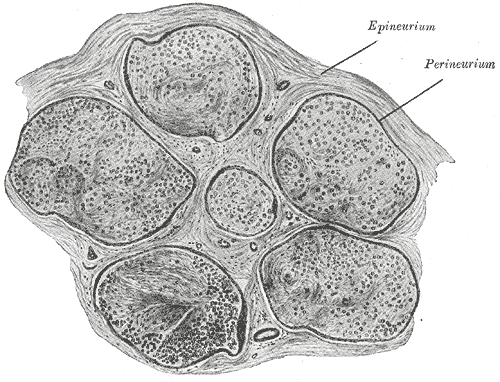
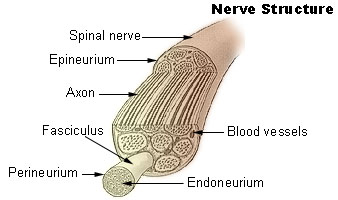
consist of myelin (in the PNS). A peripheral nerve fiber consists of an axon,
myelin sheath, Schwann cells and its endoneurium. There are no endoneurium and
Schwann cells in the central nervous system.
In myelinated axons, ionic currents are confined to the nodes of Ranvier
and far fewer ions leak across the membrane than in unmyelinated axons,
saving metabolic energy.
Myelin decreases capacitance and increases electrical resistance across the
cell membrane.
Myelin permits large organisms to exist by enabling fast communication between
distant body parts.
When a peripheral nerve fiber is severed, the myelin sheath provides a track
along which regrowth can occur. However, the myelin layer does not ensure a
perfect regeneration of the nerve fiber. Some regenerated nerve fibers do not
find the correct muscle fibers, and some damaged motor neurons of the
peripheral nervous system die without regrowth. Unmyelinated fibers and
myelinated axons of the mammalian central nervous system do not regenerate.
When an axon signal reaches a synapse, calcium channels open and calcium flows
into the cell. Vesicles, which store neutrotransmitters, then open
and release neurotransmitters into the synaptic gap. The neurotransmitters
diffuse across the synaptic gap and bind to the target cell, triggering
an action potential in the target cell.
Synapses are usually located at the terminals of axons but they can also be
located at junctions along the axon ("in passing" synapses). A single axon
with all its branches can innervate multiple parts of the brain
and generate thousands of synaptic terminals.
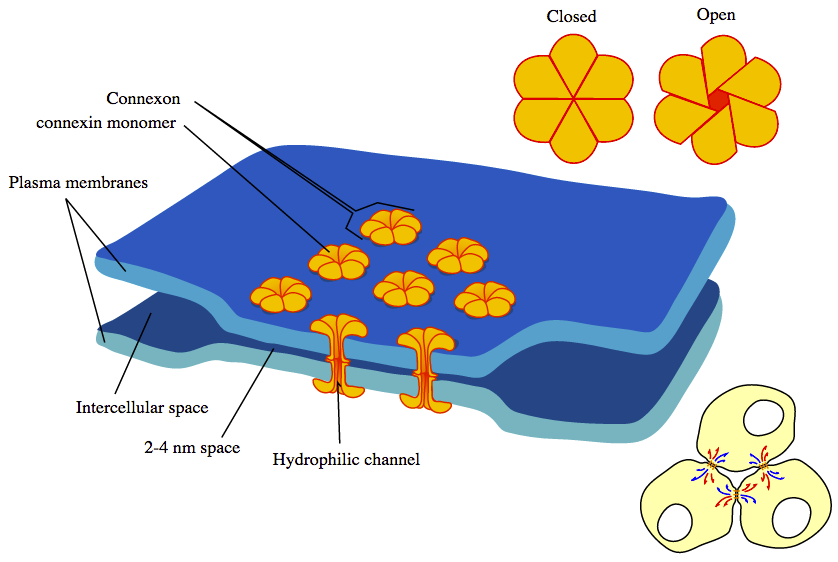
A chemical synapse can amplify signals and an electric synapse cannot.
Electric synapses are faster than chenical synapses but they can't amply
signals like chemical synapses.
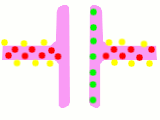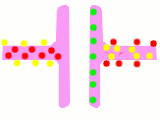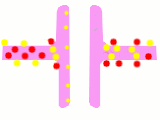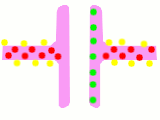
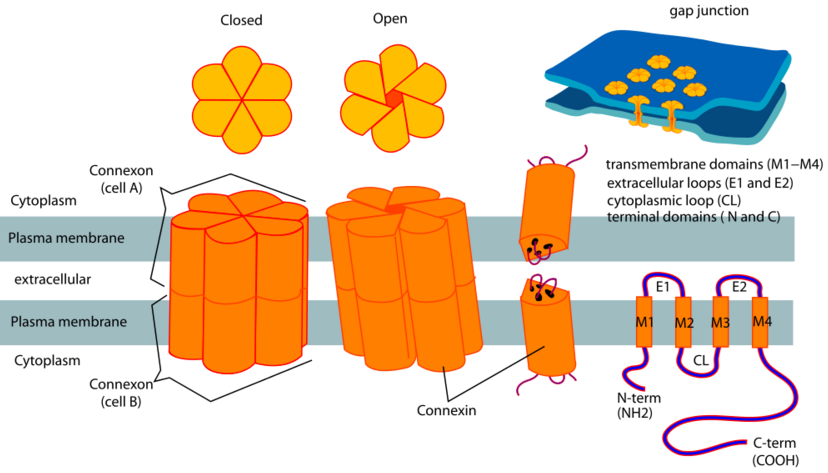
In an electric synapse, signals jump from the membrane of one cell to another
through a connexon joint. A connexon joint is a tunable iris composed of
6 connexin proteins.
The response is always the same sign as the source. For example, depolarization
of the pre-synaptic membrane always induces depolarization in the
post-synaptic membrane, and vice versa for hyperpolarization.
The response in the postsynaptic neuron is in general smaller in amplitude than
the source. The amount of attenuation of the signal is due to the membrane
resistance of the presynaptic and postsynaptic neurons.
Because electrical synapses do not involve neurotransmitters, electrical
neurotransmission is less flexible than chemical neurotransmission.
Long-term changes can be seen in electrical synapses. For example, changes in
electrical synapses in the retina are seen during light and dark adaptations of
the retina.
Surround neurons and hold them in place
Glial cells are known to be capable of mitosis whereas neurons usually are not.
In the brain "gray matter" is mostly neurons and "white matter" is mostly glial
cells.
In the cerebral cortex the distribution of glia types is:
In a supercomputer the time to multiply two numbers is much shorter than the
communication time with memory. Brains are the reverse. Signal speed is
faster than computation. A neural signal travels 20 cm (all the way across the
brain) during the time of one chemical synapse.
Human neurons are as small as physics will allow. If they were smaller then
they would be close enough for signals to jump between them even without
synapses.
Neurons do not undergo cell division. In most cases, neurons are generated by
special types of stem cells. Astrocytes are star-shaped glial cells that have
also been observed to turn into neurons by virtue of the stem cell
characteristic pluripotency. In humans, neurogenesis largely ceases during
adulthood; but in two brain areas, the hippocampus and olfactory bulb, there is
strong evidence for generation of substantial numbers of new neurons.
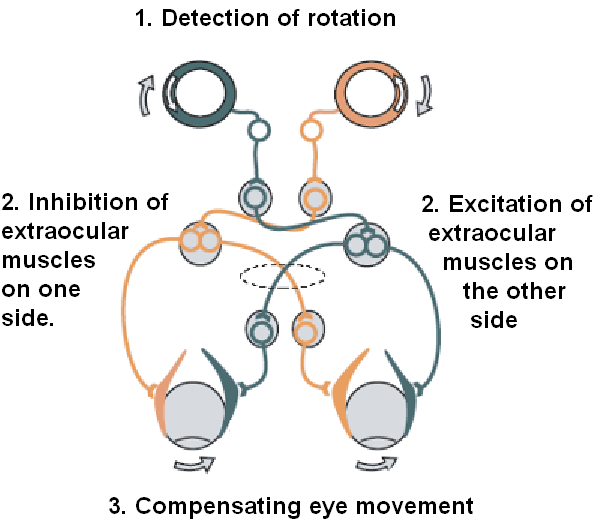
The eyes detect head movement from the vestibular system and use it to stabilize
the image.
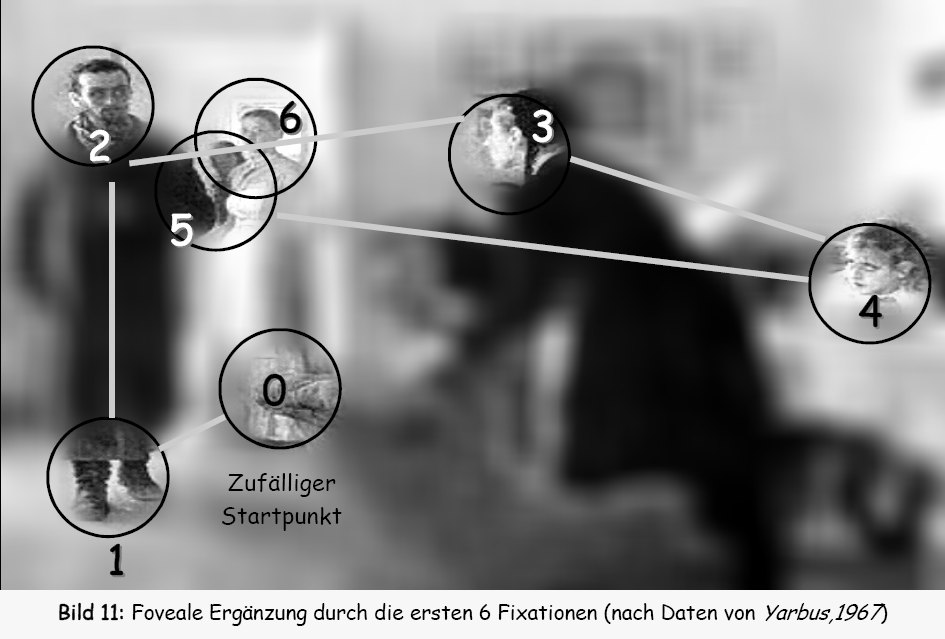
The eye moves in sudden jumps. It will be stable for an interval and then it
will make a discontinuous jump before returning to stability.
The jumps are called "saccades". Saccades are analogous to Earthquakes. When
the eye is held stable tension builds up until a saccade occurs.
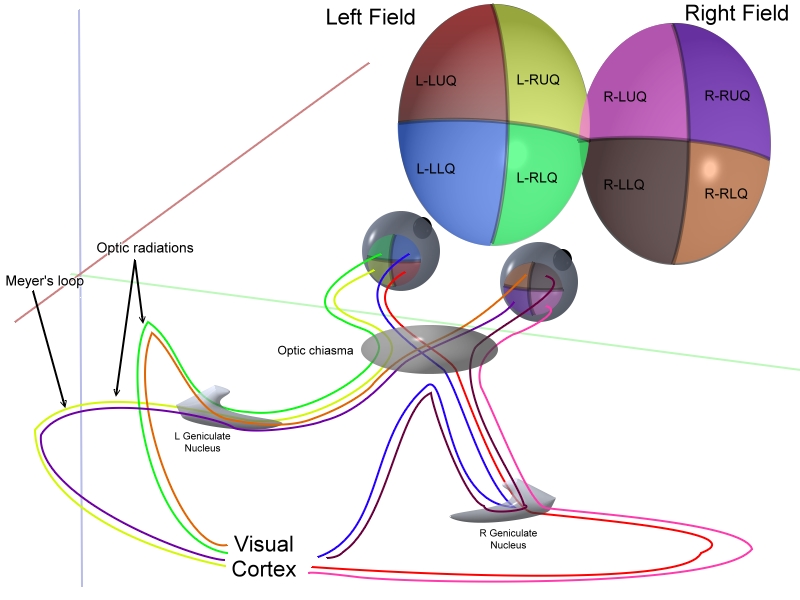
Visual information crosses at the optic chiasm before being assembled at the
rear of the brain.
Power at maximum exertion = 1500 Watts
Power used by the body at rest = 100 Watts
Power used by the brain = 20 Watts
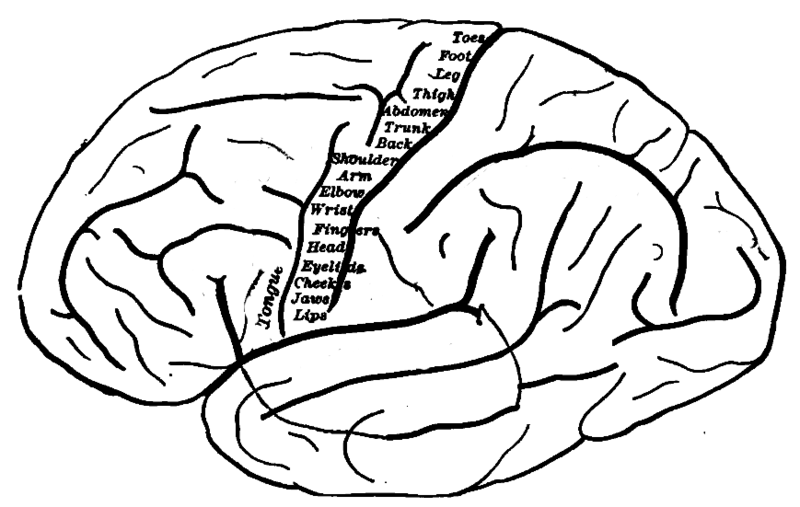
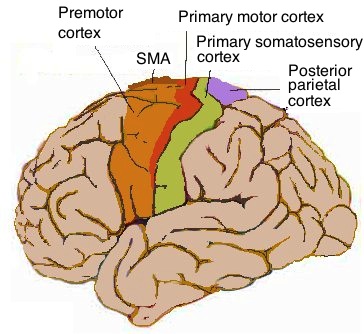
The motor cortex is in front of the somatic cortex.

Skeletal muscle cells = -95 mV
Smooth muscle cells = -60 mV
Astroglia (Glia cells) = -85 mV +- 5 mV
Neurons = -65 mV +- 5 mV
Red blood cells = -8 mV
Photoreceptor cells = -40 mV
Brain neurons = 100 billion
Brain neurons (cerebrum) = 16.3 billion
Brain neurons (cerebellum) = 69 billion
Brian glia cells = 100 billion
Brain synapes = 100 trillion
Neuron volume / Glia volume = 1.0
Neuron speed (with myelin) = 100 m/s
Neuron speed (no myelin) = 2 m/s
Axons for motor muscles = 100 m/s (16 um diameter)
Axons for sensory muscles = 10 m/s ( 8 um diameter)
Size of brain = 15 cm = 15000 neurons across
Distance between neurons = 10 μm
Axon diameter (large) = 20 μm
Axon diameter (small) = 1 μm
Membrane thickness = .0075 μm
Chemical synapse gap = .020 μm
Electric synapse gap = .0035 μm
Node of Ranvier diameter = 1.5 μm +- .5 μm
Node of Ranvier spacing =1000 μm (Distance between adjacent nodes)
Axon max size in humans = 106 μm
Dendrite max size in humans =1000 μm
Electric synapse diameter = .0016 μm
Electric synapse length = .0075 μm
Neuron body ion channels = 1 μm-2
Axon hillock ion channels = 150 μm-2
Myelin ion channels = 25 μm-2
Node of Ranvier ion channels=5000 μm-2 (Between 2000 and 12000 μm-2)
Brain neuron density = .0010 μm-3
Brain synapse density = 1.0 μm-3
Chemical synapse time = 2.0 ms
lectric synapse time = .2 ms
Sodium action potential = 1 ms (Duration)
Calcium action potential = 100 ms (Duration)
Sodium-Potassium pump time = 107 ms (Hours) (Time to reach equilibrium)
Spines per dendrite =1000
Sodium ratio = 9 (Exterior concentration / interior concentration)
Potassium ratio = 20 (Interior concentration / exterior concentration)
K+ current / Na+ current = 20 (Current across membrane in resting state)
Typical membrane potential = -.07 Volts (The cell interior is negative)
Sodium reversal potential = +.10 Volts
Potassium reversal potential= -.90 Volts
Chloride reversal potential = -.07 Volts (Same as resting potential)
Membrane breakdown voltage = .2 Volts
Breakdown field (air) = 3 Volts/μm
Breakdown field (membrane) = 27 Volts/μm
Breakdown field (vacuum) = 30 Volts/μm
Breakdown field (water) = 68 Volts/μm
Membrane capacitance = 2 uF/cm2
Max action potential rate = 100 seconds-1
Axon diam. / Nerve diam. = .6 Nerve diameter corresponds to axon plus myelin sheath
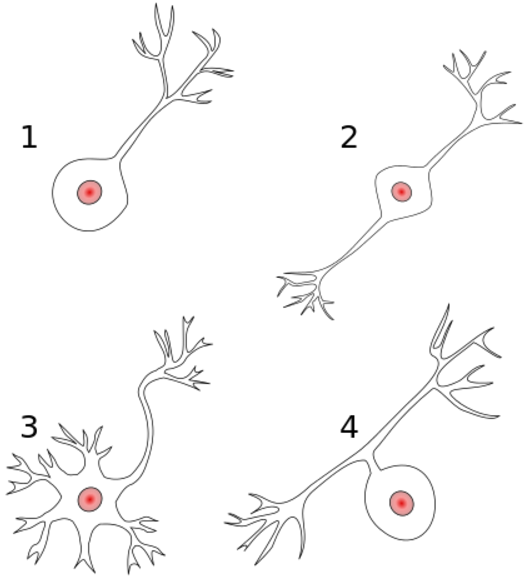
2) Bipolar neuron. Axon and dendrite on opposite ends.
3) Multipolar neuron. One axon and many dendrites.
4) Anaxonic. The axon can't be distingished from the dendrites.
Time Spacing
(ms) (nm)
Chemical synapse 2 30
Electric synapse .2 3.5

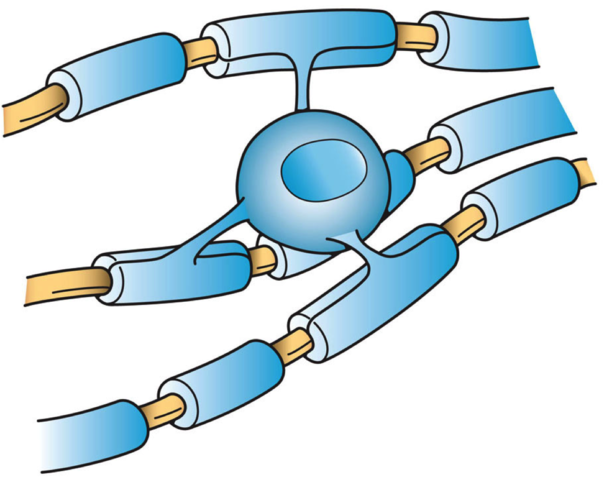
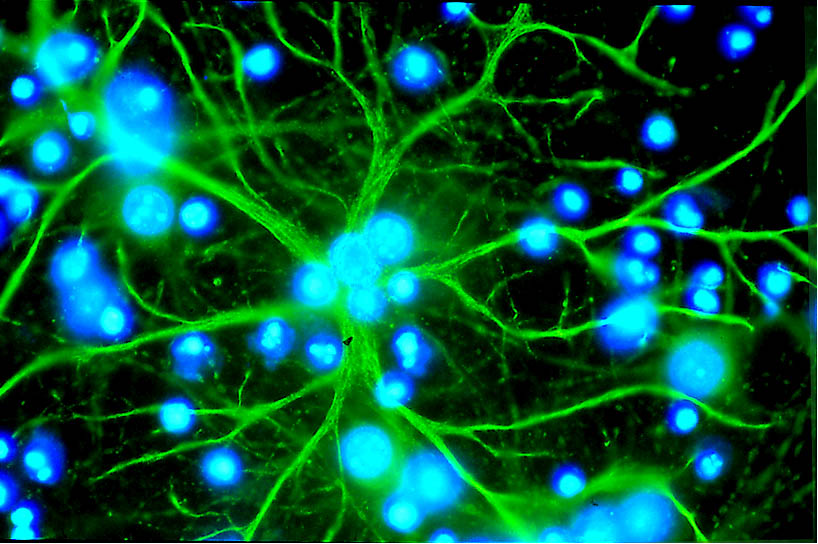
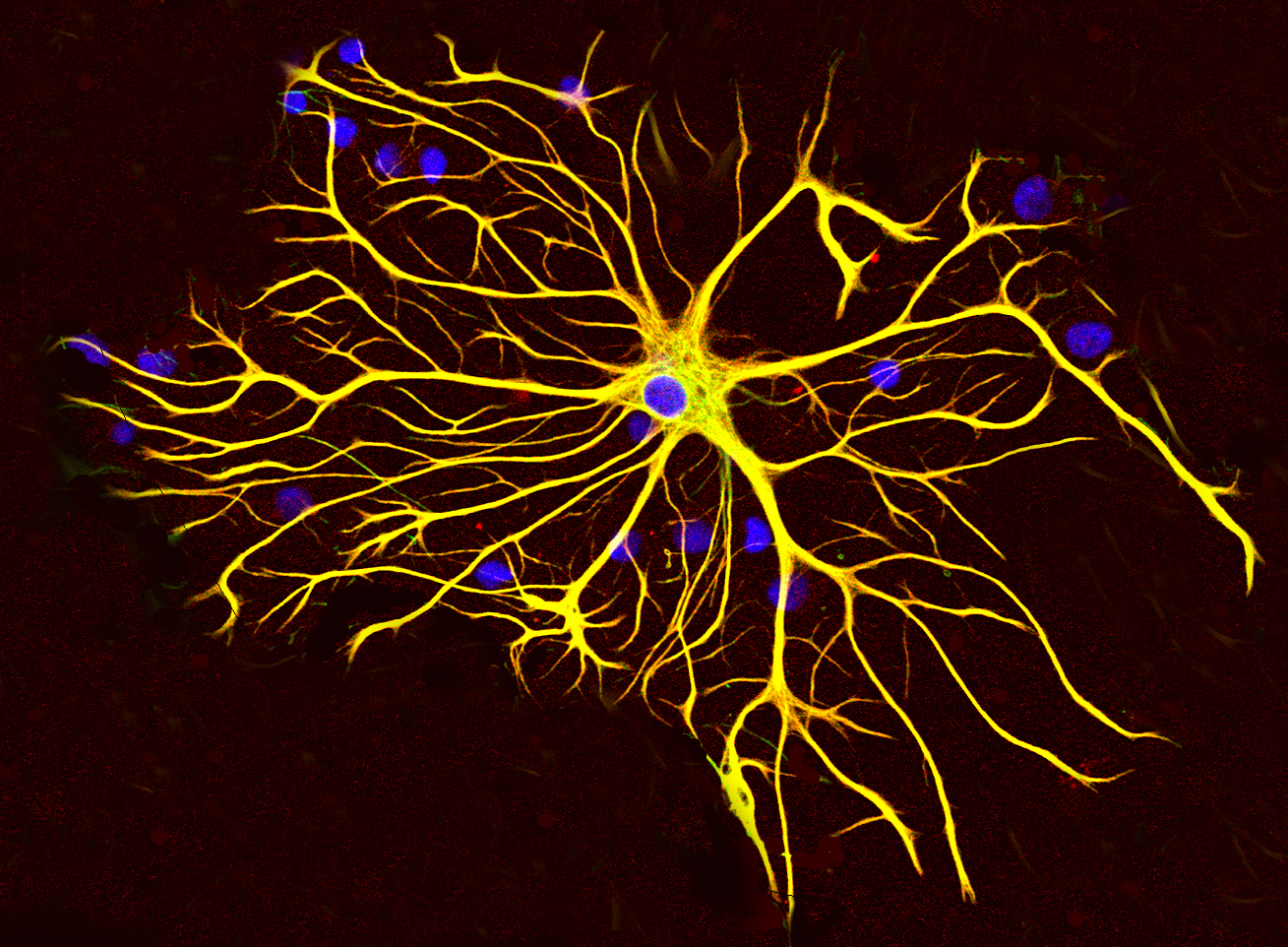
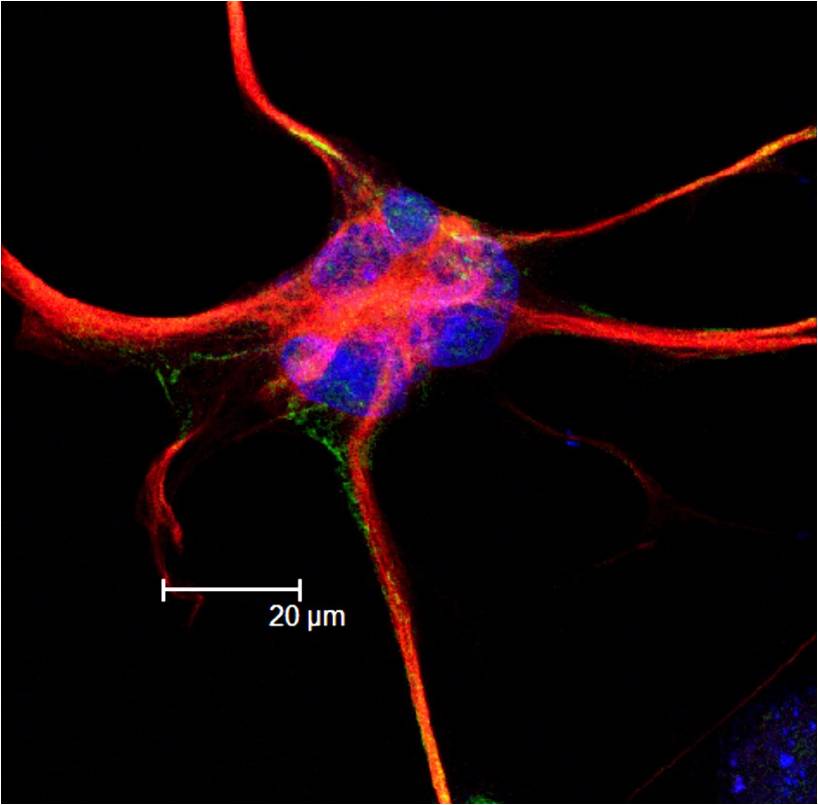
Astrocytes Provide nutrients to neurons
Microglial cell Cleanup
Oligodendrocyte Add myelin to axons in the central nervous system
Schwann cell Add myelin to axons in the peripheral nervous system
Gial cells perform functions such as:
Supply nutrients and oxygen to neurons
Supply nutrients and oxygen to neurons
Destroy pathogens and remove dead neurons
Regulate the clearance of neurotransmitters from the synaptic cleft
Release gliotransmitters such as ATP, which modulate synaptic function.
Oligodendrocytes .756
Astrocytes .173
Microglia .065
Neurons Glia
(109) (109)
Cerebral cortex 16.3 60.8
Cerebellum 69.0 16.0
CPUs Flops Devices Cycles/second Devices * Cycles/second
Brain 1 .1 1014 synapses 102 1016
Supercomputer 106 1016 106 CPUs 1010 1016
Flops = Floating point operations per second.
![]()
© Jason Maron, all rights reserved.